УДК 612.82(075.8)
Анатомия
центральной нервной системы: курс лекций для студентов психологического факультета.
Ч. II. – 2-е изд., перераб. и доп.
/ сост.: Абдуллаева Н.М., Габибов М.М., Ортабаева Л.М. – Махачкала: Издательство ДГУ, 2017. – 70 с.
Рецензенты:
Сулаквелидзе Т.С. – доктор медицинских наук, профессор, зав. кафедрой
нормальной физиологии ДГМУ;
Загирова Н.А. – кандидат медицинских наук, ассистент кафедры
нормальной физиологии ДГМУ
Издается по решению РИСа
ДГУ.
© Издательство ДГУ, 2017
Введение
Для обозначения
взаимного расположения структур нервной системы принято пользоваться специфическими
терминами, которые имеют латинское происхождение. Плоскость, проходящая вдоль
по середине тела и делящая его на правую и левую половины, называется сагиттальной. Структуры, расположенные
на спинной стороне тела, называются дорсальными,
расположенные на брюшной стороне – вентральными.
Структуры, лежащие по центру тела близко к сагиттальной плоскости, называются медиальными, лежащие вбок от
сагиттальной плоскости – латеральными.
Самые верхние точки структур называются апикальными,
лежащие в основании – базальными. Направление
к нижней части тела называется каудальным,
а к головной части – ростральным.
Тема 1. Топографическая классификация
нервной системы. Периферическая нервная система
Нервная система по топографическому принципу подразделяется
на центральную (ЦНС) и периферическую (ПНС) части. К центральной нервной
системе относятся головной и спинной мозг, состоящие из серого и белого
вещества. Серое вещество спинного и головного мозга – это скопление нервных
клеток вместе с ближайшими разветвлениями их отростков, а также ядра, или
нервные центры. Белое вещество представляет собой волокна, образованные
отростками нейронов. Нервные волокна – это проводящие пути спинного и головного
мозга, связывающие между собой как отделы ЦНС, так и различные ядра.
Периферическая
нервная система
Периферическая нервная
система – часть нервной системы, которая находится за пределами головного и спинного
мозга. К ней относятся черепные и спинномозговые нервы, нервные узлы и нервы
вегетативной нервной системы, нервные окончания, расположенные в тканях и
органах. В периферической нервной системе выделяют соматическую и вегетативную
части. Ту часть нервной системы, которая регулирует работу скелетных мышц,
называют соматической.
Посредством соматической нервной системы человек может управлять движениями,
произвольно вызывать или прекращать их. Благодаря соматической нервной системе
осуществляется связь организма с внешней средой при помощи органов чувств и
кожной чувствительности.
Часть нервной системы,
регулирующую деятельность внутренних органов (сердца, желудка, желез и т. д.),
называют вегетативной. Она
управляет вегетативными функциями организма, регулирует все обменные процессы
во всех тканях и органах. Вегетативная нервная система регулирует деятельность
органов дыхания, кровообращения, пищеварения, выделения, обмен веществ и
энергии, терморегуляцию, работу гладкой мускулатуры. Следовательно, вегетативная
нервная система регулирует функции внутренних органов, обеспечивающих нормальную
жизнедеятельность организма.
Вегетативная нервная
система морфологически и функционально подразделяется на симпатический и
парасимпатический отделы, в которых выделяют центральную и периферическую части.
Центральная часть симпатической нервной системы находится в грудном отделе
спинного мозга, а парасимпатической нервной системы располагается в стволовом
отделе головного мозга и крестцовом отделе спинного мозга. В спинном мозге
нейроны вегетативной нервной системы располагаются на границе между базальной и
крыловидной пластинами, образуя боковые рога серого вещества. Периферические
части вегетативной нервной системы состоят из нервных волокон и ганглиев,
представляющих собой скопления нервных клеток, лежащих вне пределов ЦНС.
Эфферентные волокна
центральных структур вегетативной нервной системы выходят из ЦНС в составе
смешанных черепно-мозговых волокон или по передним корешкам спинномозговых нервов.
Затем покидают общий нервный ствол и переключаются в ганглиях. Афферентные
волокна заходят в ЦНС вместе с чувствительными соматическими волокнами через
задние корешки спинного мозга или в составе черепномозговых
нервов. Ганглии в зависимости от местоположения делятся на: паравертебральные
(лежащие в непосредственной близости от позвоночника), превертебральные
(удаленные от позвоночника), экстрамуральные (лежащие
поблизости от иннервируемого органа) и интрамуральные
(расположенные непосредственно в стенке органа). Волокна, подходящие к ганглию
(преганглионарные), покрыты миелином. Волокна,
покидающие ганглий (постганглионарные), немиелинизированы и имеют серый цвет. В вегетативных
ганглиях находятся афферентные, эфферентные и ассоциативные нейроны.
В этих ганглиях количество постганглионарных волокон гораздо больше, чем количество
подходящих преганглионарных волокон. Эфферентные
вегетативные волокна оканчиваются на гладкой мускулатуре внутренних органов,
сердечной мышце и железах.
Парасимпатический и
симпатический отделы иннервируют одни и те же органы, т. е. к каждому
вегетативному органу подходят как парасимпатические, так и симпатические
окончания, часто оказывая на него противоположное действие.
Вегетативная (автономная)
нервная система обладает рядом особенностей в строении и функциях. Она
выполняет адаптационно-трофическую функцию в отношении всех тканей и органов.
Соматическая (анимальная) нервная система распространяет свое действие лишь на
скелетную мускулатуру. Таким образом, скелетная мускулатура имеет двойную иннервацию,
исходящую от обеих систем. Нейроны соматической нервной системы располагаются
более компактно, здесь нет нейронов (за исключением спинномозговых ганглиев),
вынесенных на периферию. Характерная особенность автономной нервной системы –
наличие периферических ганглиев. Нейроны соматической нервной системы
отличаются разнообразием формы и строения. Здесь встречаются гигантские
пирамидные клетки, небольшие клетки-зерна, грушевидные клетки Пуркинье в коре мозжечка и др. Нейроны вегетативной системы
в меньшей степени варьируют по величине, но имеются морфологические различия
между ними. Так, в нервных сплетениях пищеварительного тракта выявлены мультиполярные клетки Догеля 1–3
типов. Нейроны 1 типа обладают очень короткими дендритами, которые ветвятся в
пределах капсулы. Нейроны 2 типа имеют длинные дендриты, заканчивающиеся рецепторами
на периферии. У нейронов 3 типа дендриты средней величины, не выходящие за
пределы ганглия. Нейроны 1 типа являются афферентными, 2 типа – эфферентными и
3 типа – вставочными.
Крестцовый
отдел Поясничный
отдел Грудной
отдел Шейный
отдел Парасимпатическая нервная система
 Рис. 1. Вегетативная нервная
система
Рис. 1. Вегетативная нервная
система
Соматическая и вегетативные нервная системы различаются толщиной нервных
волокон. Нервные волокна автономной нервной системы имеют диаметр 1–4,5 мкм, а соматической
–
3–8 мкм. Кроме того, скорость проведения нервного импульса волокнами
соматической нервной системы равна 12–120 м/с, тогда как волокна
вегетативной нервной системы проводят импульсы со скоростью 0,3–10 м/с. Нейроны
автономной нервной системы менее возбудимы, чем соматической.
Простейшая рефлекторная дуга в обеих системах имеет трехнейронное
строение: 1-й нейрон (афферентный) располагается в спинномозговом узле; 2-й
нейрон (ассоциативный) в соматической нервной системе находится в заднем роге спинного
мозга, его аксон направляется к мотонейронам передних
рогов (3-й нейрон). В вегетативной нервной системе 2-й нейрон располагается в
боковом роге спинного мозга (интермедиолатеральное ядро), а его аксон
направляется на периферию к одному из вегетативных ганглиев. 3-й нейрон
(эфферентный) рефлекторной дуги вегетативной нервной системы вынесен на
периферию в вегетативный ганглий. Два последних нейрона соответственно называются
ганглионарным и постганглионарным.
Аксон 1-го нейрона окружен миелиновой оболочкой, а второго нейрона лишен её.
Центральные структуры симпатического
отдела вегетативной нервной системы расположены в спинном мозге. Они занимают
пространство боковых рогов серого вещества от восьмого шейного сегмента до
второго-третьего поясничного (спинномозговой центр Якобсона). Миелинизированные
аксоны этого центра выходят из передних корешков спинного мозга.
Периферическая часть симпатического
отдела состоит из двух пограничных стволов – цепочек паравертебральных
ганглиев, лежащих по краям позвоночника. Ганглии в цепочке связаны
между собой межузловыми ветвями (коннективами).
Существуют и комиссуральные связи между симметричными
ганглиями. В шейном и нижнем крестцовом отделах симпатического ствола преганглионарные нервы подходят к ганглиям не из своих сегментов
спинного мозга, а из ниже- или вышележащих сегментов через коннективы ствола. В этом случае веточки
проходят через ганглии, не переключаясь в них и оставаясь миелинизированными.
В шейном отделе
имеются три шейных узла, образовавшиеся в онтогенезе при слиянии восьми симпатических
ганглиев. Два верхних шейных узла иннервируют сонную артерию, глотку, пищевод,
слюнные и щитовидные железы и сердце. Нижний шейный узел, в свою очередь, сливается
с верхним грудным симпатическим узлом, образуя крупный звездчатый ганглий. Звездчатый
ганглий иннервирует позвоночную артерию, органы грудной полости (пищевод, трахею,
вилочковую железу, аорту) и сердечную мышцу.
В грудном отделе
имеется 10–12 ганглиев. Постганглионарные ветви
первых пяти из них направляются к плевре, сердечному, легочному и аортальному
сплетениям. Узлы с 6-го по 9-й образуют большой чревный нерв, который, пройдя в
брюшную полость, оканчивается в превертебральных
узлах самого крупного нервного сплетения брюшной полости – чревного или солнечного
сплетения. В его состав входят как симпатические, так и парасимпатические
волокна. От чревного сплетения радиально отходит множество нервов, образующих
вторичные сплетения (отсюда и название – солнечное сплетение). В узлах
солнечного сплетения оканчиваются волокна многих внутренностных нервов. От
узлов этого сплетения берут начало постганглионарные
нервы, иннервирующие почти все органы брюшной полости. Последние 2–3 узла грудного отдела своими ветвями формируют
малый чревный нерв, который также уходит к узлам солнечного сплетения.
В брюшной части
пограничного симпатического ствола имеется 4–5 поясничных узла, от которых идут
ветви к брюшной части аорты, половым органам, брыжеечному ганглию, кишечнику. В
тазовой части симпатического ствола лежат 4 крестцовых узла и один непарный копчиковый
узел. Через тазовое сплетение они иннервируют тазовые органы.
Симпатические
афферентные (чувствительные) волокна от внутренних органов, несущие информацию
от многочисленных интерорецепторов, идут в составе симпатических нервов и вступают
в спинной мозг по его задним корешкам, как и чувствительные соматические афференты. Окончания симпатических волокон выделяют в
качестве медиатора норадренилин и адреналин.
При необходимости
мобилизации ресурсов организма симпатический отдел вегетативной нервной системы
увеличивает свою активность. Под действием импульсов, приходящих по симпатическим
нервам, увеличивается частота и сила сердечных сокращений, сужается просвет кровеносных
сосудов, повышается кровяное давление, тормозится двигательная и секреторная активность
пищеварительной системы.
Высшим центром, регулирующим
работу соматических и вегетативных функций, является кора больших полушарий. В
ней имеются проекции как парасимпатических, так и симпатических нервов.
Чувствительные пути вегетативных органов проецируются в лимбическую
и ростральные части коры (орбитальная, двигательная зоны). Эти
проекции строятся на топическом принципе – рядом расположенные органы
проецируются в соседние зоны коры. Парасимпатические и симпатические проекции
одних и тех же органов проецируются в одни и те же или близко расположенные
участки коры. Однако парасимпатические проекции в коре представлены гораздо шире,
чем симпатические.
Центральные структуры парасимпатического
отдела вегетативной нервной системы расположены в стволе мозга (средний
мозг, Варолиев мост и продолговатый мозг) и в крестцовом отделе спинного мозга. Периферические части образованы
эктрамуральными и интрамуральными
ганглиями и нервами. Из среднего мозга парасимпатические ветви уходят в
составе глазодвигательного нерва (III пара). Затем преганглионарные
волокна направляются к ресничному экстрамуральному
ганглию глазницы. Постганглионарные волокна этого
ганглия иннервируют гладкую мускулатуру ресничного тела и кольцевых мышц
зрачка, т. е. являются двигательными. Варолиев мост
покидается парасимпатическими волокнами в составе лицевого нерва (VII пара). На
периферии они образуют преганглионарные веточки
нескольких экстрамуральных узлов, иннервирующих
железы слизистой оболочки носа и нёба, слезные железы, подчелюстную и подъязычную
слюнные железы. Таким образом, парасимпатические веточки лицевого нерва являются
секреторными.
Из продолговатого
мозга в составе языкоглоточного нерва (IX пара) уходят также секреторные парасимпатическое
ветви, которые направляются к ушному экстрамуральному
ганглию, иннервирующему околоушные слюнные железы и железы слизистой щек и губ.
Блуждающий нерв (X пара) является самой значительной частью парасимпатического
отдела вегетативной нервной системы. Его ветви направляются к сердечному, бронхиальному
и чревному сплетениям, а также к интрамуральным узлам
в стенках внутренних органов грудной, брюшной полостей и полости большого таза.
Парасимпатические
ветви крестцовой части спинного мозга берут начало в боковых рогах серого вещества
II–IV крестцовых
сегментов и направляются к превертебральным ганглиям
нижнего подчревного сплетения и интрамуральным
ганглиям органов малого таза. Окончаниями парасимпатического отдела вегетативной
нервной системы выделяется медиатор ацетилхолин. Парасимпатический отдел
вегетативной нервной системы регулирует работу внутренних органов в условиях
покоя. Его активация способствует снижению частоты и силы сердечных сокращений,
снижению кровяного давления, увеличению как двигательной, так и секреторной
активности пищеварительного тракта.
Филогенез. Выделение вегетативной нервной системы из общей
структуры ЦНС начинается уже у беспозвоночных. У кольчатых червей из клеток подглоточных узлов выделяются самостоятельные ганглии,
связанные с кишечной трубкой. Разделение вегетативной нервной системы на симпатическую
и парасимпатическую наблюдается уже у насекомых. Низшие позвоночные
(круглоротые) имеют блуждающий нерв и висцеральные (вегетативные) ветви
спинномозговых нервов. В различных группах позвоночных существуют свои особенности
строения вегетативной нервной системы. У млекопитающих четко проявляется тенденция
к слиянию симпатических ганглиев. Клетки ганглиев периферических отделов
вегетативной нервной системы возникают из ганглиозной пластинки.
Онтогенез. В онтогенезе парасимпатическая нервная система возникает
раньше, чем симпатическая. Сначала наблюдается формирование чувствительных
ганглиев и миграция нервных элементов на периферию по их нервам. Так, в сердце
на стадии зародыша 7,5–11 мм, уже существует интрамуральное
парасимпатическое сплетение, в то время как симпатические волокна прорастают к
нему на стадии 17–23 мм. На стадии 9–15 мм у зародыша наблюдается две пары
параллельно идущих тяжей – закладки пограничных симпатических стволов и лежащие
вентральнее их закладки превертебральных
сплетений брюшной полости. У зародыша
Тема 2. Центральная
нервная система. Спинной мозг
К центральной нервной
системе относятся те части нервной системы, тела нейронов которой защищены
позвоночником и черепом – спинной и головной мозг. Кроме того, головной и спинной
мозг защищены тремя оболочками (твердой, паутинной и мягкой). Головной мозг
анатомически делят на пять отделов: продолговатый мозг; задний мозг,
образованный Варолиевым мостом и мозжечком, средний
мозг, промежуточный мозг, образованный таламусом, эпиталамусом,
гипоталамусом и метаталамусом; конечный мозг,
состоящий из больших полушарий, покрытых корой.
Под корой
располагаются базальные ганглии. Продолговатый мозг, варолиев
мост и средний мозг являются стволовыми структурами головного мозга. Все отделы
ЦНС пронизывает полость, заполненная ликвором. В спинном и среднем мозге она
сужена, и это сужение соответственно называется центральным спинномозговым
каналом и сильвиевым водопроводом. В остальных
отделах полость образует расширения – желудочки: IV желудочек в продолговатом и
заднем мозге; III желудочек в промежуточном; боковые желудочки в больших полушариях.
Спинной мозг
представляет собой наиболее древнюю часть нервной системы. Располагается
спинной мозг в спинномозговом канале, образованном дугами позвонков, рострально он переходит в продолговатый мозг, и его верхняя
граница лежит на уровне верхнего края первого шейного позвонка. Каудально – заканчивается
на уровне I–II поясничных позвонков (у новорожденного на уровне II–III поясничных позвонков)
мозговым конусом. Спинной мозг представляет собой длинный, цилиндрической формы тяж, уплощенный в дорсовентральном направлении и изогнутый в соответствии с
кривизной позвоночника. Протяженность нервной
трубки у взрослого человека составляет 43–45 см (у мужчин) и 41–42 см (у
женщин) с почти редуцированным центральным каналом. Масса спинного мозга
колеблется в пределах 34–38 г.
В шейном и
пояснично-крестцовом отделах имеются два крупных утолщения: шейное, которое
особенно хорошо выражено на уровне V–VI шейных
невротомов, и пояснично-крестцовое, с наибольшей
шириной на уровне III–IV поясничных сегментов. Шейное утолщение соответствует выходу спинномозговых нервов,
направляющихся к верхним конечностям. Поясничное
утолщение соответствует выходу нервов, следующих к нижним конечностям. В
этих участках спинного мозга число нейронов и волокон гораздо больше, чем в
других его отделах. Внутри спинного мозга проходит центральный канал, заполненный спинномозговой жидкостью. Рострально он переходит в полость IV желудочка, каудально
слепо заканчивается конечным желудочком Краузе, расположенным в области верхней части мозгового конуса.
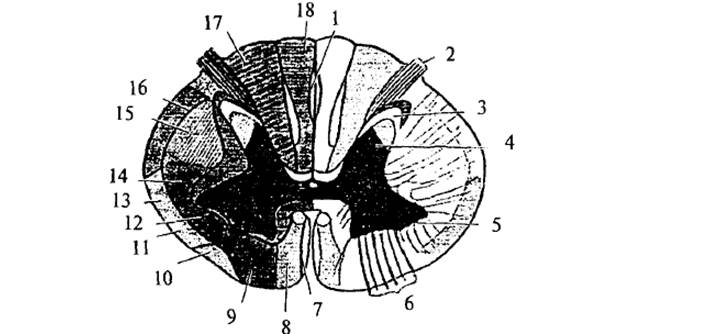
Рис. 2. Спинной мозг
(поперечный срез): 1 – овальный пучок заднего канатика; 2 – задний корешок; 3 –
субстанция Роланда; 4 – задний рог; 5 – передний рог; 6 – передний корешок; 7 –
тектоспинальный путь; 8 – вентральный кортикоспинальный путь; 9 – вентральный
вестибулоспинальный путь; 10 – оливоспинальный путь;
11- вентральный спиномозжечковый путь; 12 – латеральный вестибулоспинальный
тракт; 13 – спиноталамический тракт и тектоспинальный
тракт; 14 – руброспинальный тракт; 15 – латеральный кортикоспинальный
путь; 16 – дорсальный спиномозжечковый путь; 17 – путь Бурдаха;
18 – путь Голля
Снаружи спинной мозг
покрыт тремя оболочками, которые развиваются из мезенхимы. Мягкая, или сосудистая, оболочка содержит разветвления кровеносных сосудов, которые затем
внедряются в спинной мозг. Она имеет два слоя: внутренний, сросшийся со спинным
мозгом, и наружный. Между паутинной и мягкой оболочками находится подпаутинное пространство, заполненное цереброспинальной жидкостью.
Твердая оболочка – это длинный просторный мешок, охватывающий спинной
мозг. Рострально твердая оболочка спинного мозга срастается
с краями большого затылочного отверстия, а каудально заканчивается на уровне второго
крестцового позвонка. Твердая оболочка не примыкает к стенкам позвоночного канала,
между ними имеется эпидуральное пространство,
заполненное жировой клетчаткой и венозными синусами. Твердая оболочка связана с
паутинной в области межпозвоночных отверстий на спинномозговых узлах, а также у
мест прикрепления зубчатой связки.
Зубчатая связка
представляет собой тонкую и прочную парную пластину, которая начинается от
боковой поверхности мягкой оболочки спинного мозга, посередине между выходом
передних и задних корешков и, направляясь латерально,
разделяется на зубцы. Зубцы своими верхушками достигают и паутинной, и твердой оболочек.
Зубчатая связка, а также содержимое эпидурального, субдурального и лимфатического пространств предохраняют
спинной мозг от повреждений.
По поверхности
спинного мозга проходят продольные борозды. На вентральной поверхности спинного
мозга залегает глубокая передняя срединная щель, в которую проникает плотно
охватывающая спинной мозг мягкая мозговая оболочка. На дорсальной поверхности
располагается очень узкая задняя центральная борозда. Эти две борозды делят
спинной мозг на правую и левую половины.
По бокам от спинного
мозга отходят два ряда передних и задних корешков. Передние корешки
образованы эфферентными волокнами мотонейронов, задние
– афферентными волокнами чувствительных нейронов спинномозговых узлов.
Участок мозга с двумя
парами отходящих от него корешков называется сегментом. В спинном мозге
насчитывается 31 сегмент, каждый соответствует одному из позвонков. В шейном отделе
– 8 сегментов, в грудном – 12, в поясничном и крестцовом – по 5, в копчиковом –
1.
Так как рост спинного
мозга отстает во время онтогенетического развития от роста позвоночника, имеется
несоответствие между расположением сегментов спинного мозга и одноименными позвонками.
Поскольку нервы выходят из позвоночника через определенные межпозвоночные
отверстия, корешки удлиняются. Поэтому и направление корешков неодинаково: в
шейном отделе они отходят почти горизонтально, в грудном – спускаются косо вниз,
в пояснично-крестцовом – прямо вниз.
Ниже второго
поясничного позвонка спинномозговая полость заполнена пучком корешков,
спускающихся параллельно друг другу вниз и создающих так называемый конский
хвост. Оба корешка (передний и задний), прилегая друг к другу, направляются к
межпозвоночному отверстию и, соединяясь в области межпозвоночного отверстия,
образуют с каждой стороны смешанные спинномозговые нервы. Задний корешок у
места его соединения с передним имеет утолщение – спинномозговой узел, где располагаются
тела афферентных нейронов.
На поперечном срезе
внутри спинного мозга находится серое вещество, которое напоминает очертания буквы H или бабочку
с расправленными крыльями. Серое вещество проходит по всей длине спинного мозга
вокруг центрального канала. В каждой половине
спинного мозга оно образует два выступа – вентральный (передний) и дорсальный
(задний), называемые серыми столбами, или рогами. Передний рог более массивный, задний – более тонкий.
Правая и левая
половины серого вещества спинного мозга соединяются между собой серой спайкой.
В центре серой спайки залегает центральный канал. На некоторых сегментах
шейного и грудного отделов спинного мозга серое вещество образует, кроме
переднего и заднего серых столбов, боковой столб, или рог, расположенный на
уровне серой спайки. Нейроны серого вещества группируются в ядра, которые
вытягиваются вдоль спинного мозга и имеют вид веретен. На верхушке заднего рога
залегает желатинозная субстанция Роланда,
богатая нейроглией и большим количеством нервных клеток, которые своими отростками
связывают сегменты различных уровней друг с другом.
Между рогами
располагается центральная часть серого вещества – промежуточная зона. В промежуточной
зоне у основания заднего рога с медиальной стороны в пределах от VII шейного до
III поясничного сегментов находится группа нервных клеток, образующая
дорсальное ядро, или столб Кларка. Передние рога массивнее задних. Их образуют
довольно крупные мотонейроны, имеющие длинные аксоны,
которые образуют передние (двигательные) корешки спинного мозга. Они покидают
ЦНС в составе смешанного спинномозгового нерва и направляются к скелетной
мускулатуре.
Основную массу
нейронов спинного мозга составляют собственные нейроны, отростки которых не выходят
за пределы ЦНС. Выделяют интернейроны, или вставочные нейроны, – мелкие клетки с
короткими отростками, не покидающими серого вещества, и канатиковые, или
пучковые, клетки – более крупные клетки, отростки которых образуют белое
вещество. Серое вещество вместе с передними и задними корешками составляют сегментарный
аппарат спинного мозга, основной функцией которого является осуществление
рефлекторных реакций.
Белое вещество составляет
проводниковый аппарат спинного мозга. Белое вещество осуществляет связь
спинного мозга с вышележащими отделами ЦНС, поэтому оно развивалось параллельно
с развитием головного мозга и цефализацией. Белое вещество
залегает на периферии спинного мозга. Передняя срединная щель и задняя и
боковые борозды разделяют белое вещество каждой половины спинного мозга на так
называемые канатики. Выделяют восходящие
и нисходящие пути белого вещества спинного мозга.
Восходящие пути состоят из аксонов клеток спинальных ганглиев и
пучковых клеток серого вещества. К восходящим путям относятся:
1) тонкий (нежный)
пучок Голля;
2) клиновидный пучок Бурдаха. Эти пути сформированы из отростков клеток спинальных
ганглиев (тонкого – от 19 нижних сегментов, а клиновидного – от 12 верхних
сегментов спинного мозга). Их волокна вступают в спинной мозг через задние корешки
и отдают коллатерали нейронам серого вещества. Сами же аксоны достигают
одноименных ядер продолговатого мозга;
3) вентральный и
латеральный спиноталамические пути начинаются от чувствительных клеток
спинномозговых ганглиев, которые переключаются на пучковых клетках задних рогов
серого вещества. Аксоны этих клеток переходят по серому веществу на
противоположную сторону и достигают переключательных ядер таламуса;
4) дорсальный
спиномозжечковый путь Флексинга. Этот путь начинается
нейронами спинномозговых узлов, переключается на клетках ядра Кларка. Аксоны не
переходят на противоположную сторону и, достигая продолговатого мозга, через нижнюю
ножку мозжечка вступают в ипсилатеральную (той же стороны)
половину мозжечка;
5) вентральный
спиномозжечковый путь Говерса также начинается
нейронами спинномозговых узлов, затем переключается на клетки ядра промежуточной
зоны. Аксоны переходят на противоположную сторону и поднимаются до верхних
отделов ствола, где вновь перекрещиваются и вступают в ипсилатеральную
половину мозжечка по его верхним ножкам.
К нисходящим путям относятся:
1) латеральный и вентральный кортикоспинальные
(пирамидные) пути. Эти пути начинаются от пирамидных нейронов нижних слоев
моторной зоны коры. Они проходят через белое вещество больших полушарий, основание
ножек среднего мозга, по вентральным отделам Варолиева
моста и продолговатого мозга в спинной мозг. Латеральный путь перекрещивается в
нижней части пирамид продолговатого мозга и заканчивается на нейронах основания
заднего рога. Вентральный путь, не перекрещиваясь, пересекает пирамиды
продолговатого мозга. Перед вступлением в передний рог серого вещества
соответствующего сегмента спинного мозга волокна этого пути переходят на
противоположную сторону и заканчиваются на мотонейронах
передних рогов контрлатеральной стороны;
2) руброспинальный путь. Начинается от
красных ядер среднего мозга, перекрещивается на уровне среднего мозга. Волокна заканчиваются
на нейронах промежуточной зоны серого вещества спинного мозга;
3)
тектоспинальный путь берет начало от
клеток четверохолмия среднего мозга и достигает мотонейронов
передних рогов;
4) оливоспинальный путь. Этот путь
образован аксонами клеток олив продолговатого мозга, которые достигают мотонейронов спинного мозга;
5) вестибулоспинальные пути. Начинаются
от вестибулярных ядер продолговатого мозга и заканчиваются на клетках передних
рогов;
6) ретикулоспинальный путь. Связывает
ретикулярную формацию ствола головного мозга со спинным мозгом. Помимо этих
путей имеются внутренние, локальные пути, соединяющие
сегменты различных уровней спинного мозга между собой.
Филогенез. В
филогенезе спинного мозга отмечается вариабельность количества сегментов у представителей
различных классов и видов. Но, являясь филогенетически древней структурой, спинной
мозг мало изменяется в ходе эволюции. Пирамидные пути появляются только у млекопитающих
в связи с развитием коры больших полушарий. Удельный вес спинного мозга по
отношению к общей массе ЦНС
в ходе филогенеза уменьшается за счет увеличения массы головного мозга.
Онтогенез. В
онтогенезе спинной мозг формируется из заднего отдела медуллярной трубки. На первых
стадиях развития нервная трубка на всем протяжении состоит из трех слоев:
наружного (краевого), среднего (мантийного), внутреннего (эпендимного).
Эпендимные клетки выстилают внутренние стенки полости мозговой
трубки. Клетки мантийного слоя преобразуются в нейробласты
и спонгиобласты. Стенки трубки утолщаются неравномерно.
Основной прирост нервной ткани идет в ее боковых частях.
Нервные и глиальные
клетки боковых стенок спинномозговой трубки образуют две пластинки: крыловидную
и базальную. Крыловидная пластинка располагается дорсолатерально
вдоль дорсальной части нервной трубки, а основная, или базальная, лежит вентролатерально. Правая и левая половины пластинок соединены
с дорсальной стороны тонкой крышей, а с вентральной стороны – дном. Причем и
дно и крыша расположены в глубине около центрального канала. Отделяются
пластинки друг от друга боковыми пограничными бороздами.
Клетки крыловидной
пластинки формируют чувствительные, сенсорные, структуры задних отделов
спинного мозга (задние рога серого вещества). Базальная пластинка дает начало
клеткам передних рогов – моторным структурам. Клетки, расположенные вблизи
пограничных борозд, образуют вегетативные центры спинного мозга.
Полость нервной трубки
сужается из-за роста стенок спинного мозга и превращается в центральный спинномозговой
канал. Возникают борозды, которые делят спинной мозг на правую и левую
половины. Отростки нейронов спинного и головного мозга прорастают, покрываются
миелином и формируют белое вещество спинного мозга.
Эмбриогенез. В
начальный период внутриутробной жизни спинной мозг заполняет все пространство
позвоночного канала. Начиная с 3-го месяца зародышевой жизни, позвоночник
растет в длину быстрее, чем мозговое вещество. Поэтому только часть позвоночного
канала заполнена мозгом.
У новорожденного
спинной мозг оканчивается на уровне II–III поясничных позвонков, достигая в длину 13,6–14,8 см,
что составляет 29,5 % от длины тела. К 2 годам жизни ребенка длина спинного
мозга достигает
На поперечном срезе
вид спинного мозга ребенка такой же, как у взрослого. Однако у новорожденного
особенно хорошо выражены шейное и пояснично-крестцовое утолщения, а центральный
канал шире, чем у взрослого. Сужение просвета канала происходит главным образом
в течение первых двух лет жизни, хотя он продолжает суживаться и в более
поздние сроки, когда наблюдается увеличение массы серого и белого вещества.
Объем белого вещества увеличивается в основном за счет собственных пучков
сегментарного аппарата, который формируется раньше, чем проводящие пути,
соединяющие спинной мозг с головным.
Тема 3. Головной
мозг. Продолговатый мозг
Головной мозг с окружающими его оболочками находится в полости черепа. В
связи с этим его выпуклая верхнелатеральная поверхность по форме соответствует
внутренней вогнутой поверхности свода
черепа. Нижняя поверхность – основание мозга – имеет сложный рельеф,
соответствующий форме черепных ямок внутреннего основания черепа.
Головной мозг имеет сплющено-шарообразную форму и является наиболее
массивной частью ЦНС. Его масса составляет 1/50 массы тела. Абсолютная масса
мозга не служит показателем интеллекта человека.
По происхождению и морфологическим особенностям в головном мозге выделяют
три отдела: стволовую часть, подкорковые образования и большие полушария.
Ствол включает продолговатый мозг, Варолиев
мост, ножки мозга, крышу и средний мозг вместе с промежуточным мозгом. Мозжечок
принято выделять как малый мозг. Ствол сформировался из второго и третьего
мозговых пузырей. Подкорковая область и большие полушария развиваются из
первого мозгового пузыря.
Продолговатый мозг. В полости черепа спинной мозг без резких морфологических изменений
переходит в продолговатый мозг. Поэтому
он в дорсальной части имеет отчетливые признаки сходства со спинным мозгом.
Вследствие того, что
продолговатый мозг является продолжением спинного мозга, он сохраняет в
некоторой степени сегментарность и белое вещество,
как и в спинном мозге, занимает поверхностное положение. По нему тянутся от
спинного мозга передняя срединная щель, задняя срединная борозда, передние и
задние боковые борозды. Внутри продолговатого мозга проходит центральный канал.
Полость центрального канала на уровне продолговатого мозга расширена и образует
нижнюю часть IV желудочка мозга, имеющего ромбовидную форму.
Крыловидные пластинки
дорсальной части мозговой трубки расходятся в стороны. Крыша мозга истончена, в
ней располагается сосудистое сплетение IV желудочка. Поэтому продолговатый мозг
имеет конусовидную форму. Узкий его конец является продолжением спинного мозга,
а широкий направлен в сторону моста. Протяженность его составляет
Нижней границей
продолговатого мозга являются верхние корешки шейного отдела спинного мозга. Верхняя
граница проходит с дорсальной стороны по мозговым полоскам (волокна проводящих
путей слуховой системы), идущим поперек дна IV желудочка в самой широкой его части,
а с вентральной стороны – по нижнему краю моста.
На дорсальной
поверхности продолговатого мозга хорошо заметна полость треугольной формы – нижняя
часть ромбовидной ямки дна IV желудочка. Срединная борозда ямки каудально
(вершина треугольника) переходит в центральный спинномозговой канал. Часть
крыши мозга, прикрывающая это место (точка расхождения крыловидных пластинок),
называется задвижкой. Крышу мозга
над продолговатым мозгом образуют сосудистое сплетение IV желудочка и нижний мозговой
парус. Через три отверстия в крыше мозга – срединную апертуру, или отверстие Мажанди, и два латеральные, или отверстия Люшке, полость IV желудочка сообщается с подпаутинным пространством.
Отверстие Мажанди больше латеральных, находится в задней части
нижнего мозгового паруса и открывается в расширенную часть подпаутинного
пространства – цистерну мозжечка и спинного мозга. Отверстия Люшке расположены латерально в самой
широкой части ромбовидной ямки.
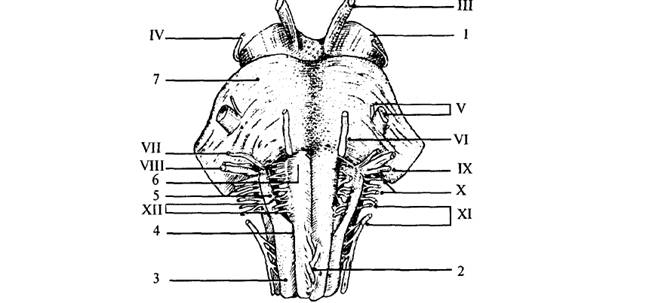
Рис. 3. Продолговатый мозг,
Варолиев мост и черепные нервы: III – глазодвигательный
нерв; IV – блоковый нерв; V – тройничный нерв; VI – отводящий нерв; VII – лицевой
нерв; VIII – преддверно-улитковый нерв (слуховой); IX – языкоглоточный нерв; X
– блуждающий нерв; XI – добавочный нерв; XII – подъязычный нерв; 1 – ножка
мозга (средний мозг); 2 – перекрест пирамид (верхний шейный отдел спинного
мозга); 3 – боковой канатик; 4 – передняя боковая борозда; 5 – олива; 6 – пирамида
продолговатого мозга; 7 – мост
Таким образом, на
уровне продолговатого мозга ликвор желудочков мозга и спинномозговая жидкость объединяются
в единую систему.
С дорсальной
поверхности продолговатого мозга от спинного мозга вдоль срединно-задней борозды
поднимаются канатики нежного пучка, латеральнее идут
клиновидные канатики. Канатики заканчиваются бугорками ядер Голля (нежного) и Бурдаха (клиновидного) на уровне нижнего угла ромбовидной
ямки. От бугорков Голля и Бурдаха рострально
идут веревчатые тела, которые являются латеральными стенками
IV желудочка. Они переходят в нижние ножки
мозжечка, имеющие вид массивных волокнистых тяжей. Нижние мозжечковые ножки
участвуют в формировании ромбовидной ямки, представляющей собой дно IV желудочка.
На вентральной
поверхности продолговатого мозга нисходящие кортикоспинальные проводящие пути
образуют расширяющиеся валики – пирамиды, лежащие по обе стороны от передней
щели. Пирамиды суживаются книзу, т. к. 2/3 их волокон переходят на
противоположную сторону, образуя перекрест пирамид, и ниже продолжаются в виде
корково-спинномозгового пути. Латеральнее от них
расположены вздутия, называемые оливами, которым внутри соответствуют
ядра серого вещества. Они отделены от пирамид передней латеральной бороздой.
Оливы образованы нейронами и на поперечном срезе напоминают складчатый мешок.
Эти образования имеют тесный контакт с мозжечком и функционально связаны с поддержанием
тела в вертикальном положении.
От продолговатого
мозга отходят IX–XII пары черепно-мозговых нервов: IX – языкоглоточный нерв, X – блуждающий нерв, XI – добавочный
нерв, XII – подъязычный нерв. Языкоглоточный нерв включает
чувствительные, двигательные и парасимпатические ветви. Он выходит из
продолговатого мозга через латеральную щель. Блуждающий нерв является
самым значительным нервом парасимпатического отдела нервной системы. Это
смешанный нерв. Добавочный и подъязычный нервы чисто двигательные
нервы. Добавочный нерв имеет веточки, выходящие из шейных сегментов спинного
мозга.
Структурная организация
продолговатого мозга отличается от спинного мозга. Скопления клеток – ядра
продолговатого мозга – перемежаются с пучками волокон, которые не всегда имеют
поверхностное расположение. Чувствительные (сенсорные) ядра черепно-мозговых нервов
располагаются дорсолатерально, двигательные
(моторные) – вентрально, вегетативные ядра занимают
промежуточное положение. Кроме ядер черепно-мозговых нервов, в продолговатом
мозге расположены ядра, на которых переключается сенсорная информация (ядра
олив, Голля и Бурдаха), и ядра ретикулярной формации,
а также дыхательный, сосудодвигательный и пищеварительные центры.
В продолговатом мозге
и других стволовых отделах мозга расположена особая клеточная структура – ретикулярная
формация. Она образована переплетением нервных волокон и лежащих между ними
нейронами и их скоплениями. По своему виду они напоминают мелкие ядра.
Ретикулярная формация
является продолжением аналогичной структуры спинного мозга. Она занимает
обширное пространство между трактами и ядрами ствола мозга. Ретикулярная
формация – слабо дифференцированная масса нервных клеток, регулирующая
состояние коры большого мозга.
В функциональном
отношении ретикулярная формация ствола представляет собой единую структуру, в
которой выделено до 98 ядерных групп. Эта структура связана со всеми органами
чувств, двигательными и чувствительными областями коры большого мозга,
зрительными буграми, подбугровой областью и спинным мозгом.
Основными ядрами
являются: ядра шва и гигантоклеточное ядро продолговатого мозга, центральное и
ретикулярное ядра моста. Клетки ретикулярной формации очень разнообразны по форме
и величине. Для них характерно наличие значительно разветвленного дендритного
дерева и длинных аксонов.
Афферентные входы ретикулярная
формация получает как по коллатералям восходящих (сенсорных) путей, так и от вышележащих
структур, в том числе от коры больших полушарий и мозжечка.
Таким образом, ретикулярная
формация интегрирует влияние большого числа мозговых структур. В свою очередь сама
она оказывает влияние как на вышележащие, так и на нижележащие структуры.
Нисходящие и
восходящие волокна ретикулярной формации покидают ядра на всем ее протяжении,
не имея четких пространственных разграничений. Кроме того, существуют аксоны, которые
Т-образно ветвятся. Одна из ветвей идет вниз, а другая – в верхние отделы
головного мозга. Нисходящие волокна образуют ретикулоспинальный тракт. Через
него ретикулярная формация оказывает влияние как на двигательную деятельность
спинного мозга (осуществление спинальных рефлексов), так и на вегетативную
регуляцию (сосудодвигательная, дыхательная, пищеварительная функции).
Ретикулярная формация
воздействует на соматические и вегетативные центры в двух противоположных
направлениях – торможения и возбуждения. Восходящие влияния ретикулярной
формации направлены на регуляцию деятельности коры больших полушарий.
Большинство ретикулярных волокон достигает коры через переключение в неспецифических
ядрах таламуса. Значительная часть нейронов ретикулярной формации связаны синапсами
с двумя или тремя афферентными волокнами.
Действие восходящих
ретикулярных влияний заключается в широкой активации корковых структур.
Торможение активности ретикулярной формации ведет к наступлению сна, активация
ее приводит к реакции пробуждения.
Характерной
особенностью нейронов ретикулярной формации является их высокая чувствительность
к химическим факторам. Такие факторы, как уровень углекислого газа, кислорода, адреналина,
ацетилхолина и серотонина, относительно небольшие концентрации
фармакологических веществ, изменяют активность нейронов ретикулярной формации,
а вместе с этим и ее влияние на кору больших полушарий, соматические и вегетативные
рефлексы.
Филогенез. В филогенезе развитие продолговатого мозга
определяется развитием чувствительных органов боковой линии, слуха, тактильной
чувствительности. У всех позвоночных животных, кроме млекопитающих, в
продолговатом мозге расположены ядра V–XII пар нервов. У низших позвоночных
пирамиды отсутствуют, а у млекопитающих они ясно различимы. Оливы появляются
только у высших обезьян.
Поскольку наибольший
кортикальный контроль над двигательными структурами наблюдается у человека, у
него же наиболее выражены и пирамиды, и оливы.
Филогенетически ретикулярная
формация является древней структурой. У низших позвоночных она распространена
по всей ЦНС. У высших позвоночных эта структура хорошо представлена только в
стволовых отделах мозга, так как функцию регуляции берут на себя конечный мозг
и мозжечок.
Онтогенез. В
онтогенезе продолговатый мозг является производным ромбовидного мозгового пузыря,
который делится на задний и добавочный мозг. Полость ромбовидного мозга не делится
и остается общей для продолговатого и заднего мозга – IV желудочком. Добавочный мозг развивается в
продолговатый. Продолговатый мозг формируется так же, как и спинной.
В структурах
продолговатого мозга формируются ядра черепно-мозговых нервов. Причем почти все
ядра нервов (тройничного, отводящего, лицевого, слухового), принадлежащие во
взрослом состоянии мосту, закладываются первоначально в ростральных отделах
продолговатого мозга, и только позже они перемещаются в структуры заднего
мозга.
Сосудистое сплетение
IV желудочка на начальных этапах эмбрионального развития человека представляет
собой сплошную пластинку. В дальнейшем она прорывается, образуя отверстия: места
сообщения между полостью IV желудочка и полостью подпаутинного
пространства.
Тема 4. Задний мозг (Варолиев
мост. Мозжечок)
Варолиев мост и мозжечок принято объединять в единую структуру
под общим названием задний мозг. Задний
мозг развивается из заднемозгового пузыря,
являющегося производным ромбовидного пузыря. Вентральная часть заднего мозга
представляет собой продолжение стволовых структур и носит название Варолиевого моста.
Варолиев мост – это вентральная часть заднего мозга. Он образует
структуры ростральной части дна IV желудочка, являющегося производным
ромбовидного пузыря зародыша и поэтому его ограничивают продолговатый мозг,
мост и мозжечок. Дорсальная поверхность
моста представляет собой верхний треугольник ромбовидной ямки. Полость
ромбовидной ямки рострально сужается и переходит в водопровод
среднего мозга. Сверху полость ромбовидной ямки прикрыта верхним мозговым парусом,
который вместе с нижним мозговым парусом и сосудистым сплетением образует крышу
IV желудочка, имеющую вид шатра. Латеральные стенки IV желудочка в области
моста образованы средними и верхними ножками мозжечка.
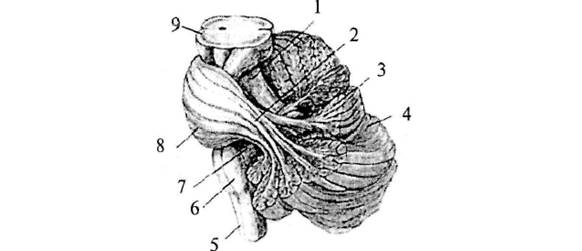
Рис. 4. Варолиев
мост и мозжечок: 1 – верхняя ножка мозжечка; 2 – средняя ножка мозжечка; 3 – лист
(долька мозжечка); 4 – левое полушарие мозжечка; 5 – спинной мозг; 6 – продолговатый
мозг; 7 – нижняя ножка мозжечка; 8 – мост; 9 – средний мозг
Вентральная
поверхность Варолиева моста представляет собой мощную
поперечно-волокнистую выпуклость белого вещества. По центру вентральной
поверхности моста проходит глубокая борозда – канавка основной артерии мозга. Латерально вентральная выпуклость переходит в мощные
средние ножки мозжечка. От Варолиевого моста отходят четыре пары черепно-мозговых
нервов. V – тройничный нерв; VI – отводящий нерв; VII – лицевой нерв;
VIII – преддверно-улитковый, или слуховой, нерв.
На поперечных срезах,
как и в продолговатом мозге, в нем выделяются белое вещество и ядра серого вещества.
Поперечные волокна, составляющие трапециевидное тело, делят толщу моста на
более крупную вентральную (основание моста) и дорсальную (покрышка моста)
части. В вентральной части преобладает белое вещество проводящих путей,
являющихся продолжением проводящих путей ножек среднего мозга. Серое вещество
вентральной части моста образует собственные ядра моста (ядра основания моста).
В этих ядрах заканчиваются волокна, идущие из коры больших полушарий: нисходящие
кортикомостовые пути и коллатерали от кортикоспинальных путей.
От собственных ядер
моста отходят волокна, которые переходят на противоположную сторону и образуют трапециевидные
тела, переходящие в средние ножки мозжечка. Дорсальная часть моста является непосредственным продолжением
продолговатого мозга. В ней располагаются переключательные ядра сенсорных систем,
ядра черепно-мозговых нервов и ретикулярной формации.
В филогенезе у низших
позвоночных животных Варолиев мост не отделен четко
от продолговатого мозга. Он обособляется только у млекопитающих. Это происходит
с развитием коры и нисходящих из нее проекционных путей. Вместе с этим растет
количество собственных ядер вентральной части моста. Это обусловливает
появление и развитие средних ножек мозжечка и его полушарий. Особенно
значительно вентральные части моста и средние ножки мозжечка выражены у
человека.
В онтогенезе мост, как
отдел заднего мозга, происходит из ромбовидного мозгового пузыря. На стадии
пяти мозговых пузырей ромбовидный мозг делится на добавочный, из которого развивается
продолговатый мозг, и задний мозг. Крыша заднего мозга преобразуется в
мозжечок, а дно и стенки его становятся структурами моста. Полость ромбовидного
мозга остается общей для продолговатого мозга и моста и является полостью IV желудочка.
Почти все ядра
черепно-мозговых нервов моста закладываются в ростральных отделах продолговатого
мозга. Их перемещение в мост происходит после образования мозгового изгиба.
На 7-й неделе развития
эмбриона клетки крыловидной пластинки продолговатого мозга мигрируют в ростровентральном направлении и образуют на вентральной
поверхности моста понтобульбарное тело, которое позже
превращается в собственные ядра моста.
Мозжечок
располагается на дорсальной поверхности ствола мозга. Он занимает заднюю черепную
ямку. Вентральная поверхность мозжечка принадлежит к парусам IV желудочка и
тесно связана со стволовыми структурами тремя парами мозжечковых ножек: с
продолговатым мозгом нижними ножками, с мостом средними ножками и со средним
мозгом – верхними ножками. Наиболее мощными являются средние ножки.
Все ножки выходят из
мозжечка рядом, а потом верхние ножки направляются к среднему мозгу вместе с
верхним мозговым парусом, а нижние – к продолговатому мозгу вместе с нижним
мозговым парусом. Рострально над мозжечком лежат затылочные
доли большого мозга, которые заходят за его дорсальные границы.
Мозжечок отделен от
большого мозга глубокой поперечной щелью. Он, как и большой мозг, покрыт тремя
оболочками. У взрослого человека масса мозжечка составляет 120–160 г.
Анатомически мозжечок
человека состоит из трех основных частей: двух полушарий и соединяющей их
средней части – червя. Поверхность мозжечка изрезана глубокими ветвящимися бороздами.
Глубокие борозды мозжечка делят полушария и червь на дольки, которые
объединяются в доли: верхние, задние и нижние. Доли отделяются друг от друга
щелями.
Между обоими
полушариями мозжечка располагается изрезанная узкими, параллельно идущими бороздами
средняя часть – червь. На нем различают верхнюю поверхность – верхний
червячок, и нижнюю – нижний червячок. Две продольно идущие бороздки на каждой поверхности
мозжечка отделяют верхний и нижний червячки от полушарий мозжечка. На верхнем и
нижнем червячках различают доли, которые состоят из нескольких извилин. Червь
оказывает регулирующее влияние на двигательную активность осевых структур
(туловище, шея и голова), а полушария связаны с работой конечностей.
Верхний червячок спереди назад состоит из следующих долей: 1) язычок
мозжечка; 2) центральная долька;
3) бугор; 4) лист
червя в виде очень узкой дольки располагающейся сзади, на границе перехода
верхнего червячка в нижний.
На нижнем червячке
в направлении спереди назад различают следующие доли: 1) узелок, к передним
отделам которого присоединяется задний мозговой парус; 2) втулочка
червя; 3) пирамида червя.
На верхней
поверхности полушарий мозжечка отмечают следующие борозды и дольки. Четырехугольная
долька разделяется передней верхней бороздой на переднюю и заднюю части.
Четырехугольная долька ограничивается задней верхней бороздой от верхней полулунной
дольки. Спереди от четырехугольной дольки залегают небольшие извилины,
так называемые крылья центральной дольки. Снизу и спереди от последних лежат небольшие
участки мозжечка – связи язычка.
На нижней поверхности полушарий мозжечка
располагаются борозды и дольки. Группа концентрически расположенных
извилин образует миндалину. Снаружи и сзади от миндалины залегает двубрюшная
долька, которая соответствует пирамиде червячка. Снаружи от миндалины и спереди
от двубрюшной дольки располагается наиболее выступающая спереди часть нижней поверхности
мозжечка – клочок. Червь и клочок являются филогенетически древними участками
мозжечка. Сзади от двубрюшной дольки лежит нижняя полулунная долька.
Нейронная организация
мозжечка существенно отличается от таковой стволовых структур. Основная масса
нейронов сосредоточена на поверхности и создает кору мозжечка, толщина которой
составляет 1–2,5 мм. Площадь ее велика, так как кора имеется и на
боковых поверхностях борозд (около 80 %). При том что масса мозжечка составляет
лишь 1/9 массы обоих больших полушарий, площадь поверхности его коры равна
площади поверхности одного из них.
Серое вещество коры, расположенное
на поверхности ветвящихся борозд, пронизывает, как дерево, белое вещество. Поэтому
рисунок, образуемый серым и белым веществом на срезах мозжечка, называют «древом
жизни» мозжечка. В глубине белого вещества имеются скопления серого вещества – парные
ядра мозжечка. B черве по обеим сторонам от средней линии имеются два ядра
шатра. Латеральнее ядер шатра в полушариях мозжечка располагаются
шаровидные ядра. Латеральнее последних в полушариях находятся
пробковидные ядра, а еще дальше – самые крупные зубчатые ядра полушарий. Последние
имеют вид волнообразно изгибающейся пластинки серого вещества.
Кора мозжечка четко
разделена на три слоя:
1) наружный – молекулярный
слой. В нем
находятся аксоны и дендриты клеток нижележащих слоев, а также звездчатые и корзинчатые клетки. В этом слое встречаются и клетки
Лугано, функция которых не установлена;
2) средний – ганглиозный
слой. Образован
крупными грушевидными клетками Пуркинье, имеющими
каждый мощное, сильно ветвящееся дендритное дерево в молекулярном слое. Эти
клетки расположены в один ряд. Количество их у человека приближается к 15 млн;
3) внутренний – зернистый
слой. Аксоны клеток-зерен направляются в молекулярный слой, где Т-образно
ветвятся и вступают в синаптические контакты с
дендритами клеток Пуркинье, корзинчатых
и звездчатых клеток.
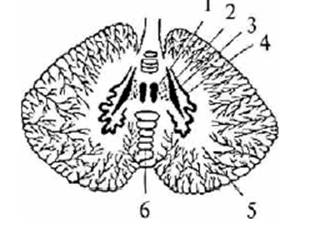
Рис. 5. Ядра мозжечка: 1 – ядро
шатра; 2 – шаровидное ядро; 3 – пробковидное ядро; 4 – зубчатое ядро; 5 – полушария
мозжечка; 6 – червь мозжечка
Дендритное дерево
клеток Пуркинье располагается в плоскости,
перпендикулярной оси борозды, а аксоны клеток-зерен – параллельно ей. На одну
клетку Пуркинье приходится около 5 тыс. клеток-зерен.
Общее количество зерновидных нейронов достигает у человека 1010–1011.
Клетки Пуркинье интегрируют информацию, приходящую из
спинного мозга и коры больших полушарий.
На соме и дендритах
клеток Пуркинье заканчиваются еще и аксоны звездчатых
и корзинчатых клеток, а также так называемых лазающих
волокон от ядер оливы (пришедших к коре мозжечка по оливо-мозжечковым
путям). Остальные афферентные пути заканчиваются в коре мозжечка в виде моховидных
волокон на клетках-зернах, а также на звездчатых и корзинчатых
клетках. Эфферентные выходы из коры
мозжечка созданы аксонами клеток Пуркинье,
заканчивающимися на клетках подкорковых ядер мозжечка. Из аксонов клеток ядер мозжечка
состоят эфференты мозжечка, связывающие его с другими
отделами ЦНС.
Афферентные и эфферентные
волокна образуют в совокупности три пары мозжечковых ножек. Через нижнюю пару
ножек мозжечок получает афференты от дорсального спиномозжечкового
пути Флексига. Здесь проходят оливо-мозжечковый
путь, пути от вестибулярных ядер VIII пары нервов и ядер V, VII, IX и X пар
черепно-мозговых нервов, а также от ядер Голля и Бурдаха
продолговатого мозга.
Через нижние ножки
выходит лишь один эфферентный путь от ядер шатра к вестибулярным ядрам
продолговатого мозга. Средние ножки имеют только афферентные волокна, идущие от
собственных ядер моста, а также коллатерали от кортикоспинальных путей. Через
эти ножки различные отделы коры больших полушарий (лобные, височные и
затылочные) связаны с мозжечком, так как на собственных ядрах моста заканчиваются
корково-мостовые нисходящие пути.
Через верхние ножки
мозжечок получает афферентные волокна от вентрального спиномозжечкового пути Говерса, а также от передних бугров четверохолмия. Основную
же массу передних ножек составляют эфферентные волокна, идущие к красному ядру, ретикулярным ядрам и буграм четверохолмия
среднего мозга, к таламическим и гипоталамическим ядрам промежуточного мозга.
Через таламические
ядра мозжечок связан с корой больших полушарий, а через красные ядра, ядра
ретикулярной формации и вестибулярные ядра – со спинным мозгом. Мозжечок
является важным участком вегетативной нервной системы.
Филогенез. Все позвоночные животные обладают мозжечком, развитие которого связано с
характером и сложностью выполняемых ими движений. Поэтому у рыб мозжечок
развивается так, что становится высшей интегративной структурой. Мозжечок рыб
состоит из тела и двух небольших возвышений. Их называют древним мозжечком, а
тело – старым мозжечком.
При выходе животных на
сушу мозжечок вначале редуцируется (у амфибий). Его развитие шло по пути
совершенствования связей спинного мозга с мозжечком и ослабления связей с вестибулярной
системой. У высших рептилий (крокодилов) и птиц формируется кора мозжечка,
имеющая два слоя клеток: гранулярный и молекулярный (с клетками Пуркинье). Тело мозжечка делится на три доли: переднюю,
среднюю и заднюю.
У млекопитающих в
мозжечке возникают структуры, тесно связанные с корой больших полушарий – новый
мозжечок. Наибольшего развития достигает зубчатое ядро. Развитый мозжечок
высших млекопитающих состоит из трех частей: древний контролирует вестибулярную
функцию (его каудальные отделы); старый связан со спинным мозгом (передняя
доля) и новый – с корой больших полушарий.
Онтогенез. В онтогенезе мозжечок развивается из мозжечковой
пластинки, являющейся крышей заднемозгового пузыря.
Полушария мозжечка образуются из боковых частей, а из средней части этой
пластинки формируется червь.
У новорожденного
мозжечок развит слабо, особенно его полушария. Извилины и борозды едва
намечены. Наиболее хорошо из всех структур развит червь. Масса мозжечка
новорожденного достигает всего 20 г, что составляет 5,4 % от массы тела. У
взрослого человека этот показатель составляет 10 %. В течение первых пяти
месяцев постнатальной жизни масса мозжечка увеличивается в 3 раза по сравнению
с периодом новорожденности. Наиболее интенсивно развиваются полушария мозжечка.
Тема 5. Средний мозг. Промежуточный мозг
Средний мозг
является верхней частью мозгового ствола. В отличие от других отделов головного
мозга он устроен менее сложно и в процессе эволюции претерпел незначительные
изменения. Его развитие связано со зрительными и слуховыми сенсорными системами.
Средний мозг делят на дорсальную
часть – крышу мозга и вентральную – ножки мозга. Крыша мозга представлена
пластинкой четверохолмия. Полость среднего мозга – это узкий канал – Сильвиев водопровод, который соединяет III и
IV мозговые желудочки. Протяженность этого узкого канала составляет 2 см. Крыша
среднего мозга, или пластинка четверохолмия, образована двумя верхними и
двумя нижними холмиками. От каждой пары холмиков в направлении промежуточного
мозга отходят проводящие пути – пары ручек холмиков. Ручки верхних холмиков заканчиваются
в латеральных коленчатых телах, а нижних – в медиальных коленчатых телах промежуточного
мозга.
У основания мозга,
впереди моста лежат ножки мозга – два симметричных толстых расходящихся
валика, которые упираются в большие полушария. Между ножками имеется
межножковая ямка, закрытая задним продырявленным пространством. На медиальную
поверхность каждой ножки выходят волокна III пары глазодвигательного нерва.
Волокна IV пары блокового нерва отходят от дорсальной поверхности среднего
мозга. Оба нерва среднего мозга являются двигательными.
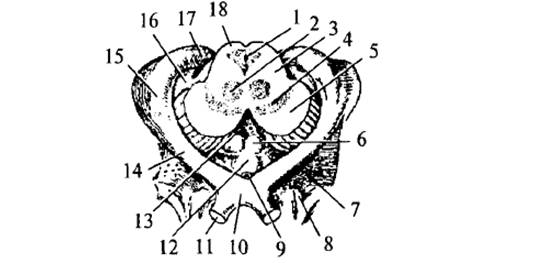
Рис.
6. Средний и промежуточный мозг: 1 – Сильвиев водопровод; 2 – красное ядро; 3 – покрышка
среднего мозга; 4 – черная субстанция; 5 – ножки среднего мозга; 6 -сосцевидные
тела; 7 – переднее продырявленное пространство; 8 – обонятельный треугольник; 9
– воронка; 10 – зрительный перекрест; 11 – зрительный нерв; 12 – серый бугор;
13 – заднее продырявленное пространство; 14 – зрительный тракт; 15 – латеральное
коленчатое тело; 16 – медиальное коленчатое тело; 17 – подушка таламуса; 18 – крыша
среднего мозга
На поперечном срезе
среднего мозга выделяют три отдела: крышу среднего мозга, покрышку, основание
ножек мозга. Наружная поверхность крыши среднего мозга покрыта тонким слоем белого вещества, переходящим
в ручки холмиков. Под этим слоем находятся ядра верхних и нижних бугров четверохолмия. Ядра верхних
бугров имеют слоистое строение. К ним приходят афферентные волокна от
зрительного тракта, от спинного мозга по спинотектальным
путям, а также коллатерали от латеральной и медиальной петель.
Отходят эфферентные
волокна к двигательным ядрам ствола и спинного мозга по тектобульбарным
и тектоспинальным путям. Верхними ручками передние бугры связаны с латеральными
коленчатыми телами. В ядрах нижних бугров заканчивается часть волокон латеральной
петли. Эфферентными же волокнами они сплетаются с медиальными коленчатыми
телами (по нижним ручкам), а также со спинным мозгом и мозговым стволом (по
тектоспинальным и тектобульбарным путям).
Верхнее двухолмие
является подкорковым зрительным центром, а нижнее двухолмие служит местом
переключения слуховых путей и играет роль слухового подкоркового центра.
В покрышке среднего
мозга имеются красные ядра, которые дают начало руброспинальному пути. В
красных ядрах заканчиваются волокна верхних ножек мозжечка. Вокруг Сильвиева водопровода располагается центральное серое
вещество. В нем находятся ядра ретикулярной формации среднего мозга, получающие
коллатерали от проходящих здесь восходящих и нисходящих путей, а свои длинные аксоны
направляющие к другим мозговым структурам и к коре больших полушарий.
Ядра блокового нерва
(IV пара) лежат в центральной части серого вещества, непосредственно у Сильвиева водопровода, на уровне нижних бугров четверохолмия.
Под дном водопровода, на уровне верхних бугров четверохолмия, находятся ядра
глазодвигательных нервов (III пара). Латерально и
кверху от красных ядер находятся слои медиальных петель, идущих от покрышки
моста. Между покрышкой и основанием ножек располагается ядро, состоящее из
клеток, богатых меланином, которое носит название черная субстанция. Основание
ножек мозга лишено ядер и образовано нисходящими из коры больших полушарий корково-спинномозговыми,
корково-мостовыми путями.
Средний мозг является
первичным зрительным и слуховым центром, осуществляющим быстрые рефлекторные
реакции (оборонительные и ориентировочные). Кроме того, красные ядра и черная субстанция
являются ядрами, контролирующими тонус скелетной мускулатуры и движения. Деятельность
красных ядер находится под значительным контролем коры больших полушарий,
влияния которой на двигательные реакции передаются по рубро
– спинномозговому пути. Поэтому красные ядра оказываются важным звеном
экстрапирамидной системы.
Филогенез. У круглоротых средний мозг представлен крышей, имеющей
связи с продолговатым мозгом. У ганоидных и костистых рыб развивается покрышка,
где формируются ядра черепномозговых нервов,
появляются связи с мозжечком. У амфибий развита крыша, представленная двухолмием.
У рептилий возникает красное ядро и формируется руброспинальный путь. У
млекопитающих появляется черная субстанция, крыша из двухолмия превращается в четверохолмие
с сохранением зрительной функции за передним двухолмием и формированием слуховой
функции в связи с задним двухолмием.
Кроме того,
устанавливаются связи среднего мозга с таламусом, базальными ганглиями и корой
больших полушарий. В центральной части среднего мозга развивается массивная
ретикулярная формация с множественными восходящими и нисходящими путями.
Онтогенез. В
онтогенезе средний мозг формируется из среднего мозгового пузыря, стенки которого
равномерно утолщаются, превращая его полость в узкий канал – водопровод.
Промежуточный мозг расположен над мозолистым телом и скрыт большими полушариями. Он каудально соединяется со средним мозгом, а рострально переходит в большие полушария конечного мозга.
Полость промежуточного
мозга представляет собой вертикальную щель, расположенную в серединной
сагиттальной плоскости, это III мозговой желудочек. Сзади эта полость переходит
в водопровод среднего мозга, а впереди соединяется с двумя боковыми желудочками
больших полушарий посредством двух межжелудочковых отверстий Монро.
Боковые
стенки III желудочка образованы медиальными поверхностями правого и левого таламусов,
дно – гипоталамусом и субталамусом. Передняя граница
подходит к нисходящим колоннам свода ниже к передней мозговой комиссуре и далее
к конечной пластинке. Задняя стенка состоит из задней комиссуры над входом в
водопровод мозга. Крыша III желудочка состоит из эпителиальной пластинки. Над
ней располагается сосудистое сплетение. Выше сплетения проходит свод, а еще
выше – мозолистое тело.
По боковым стенкам III
желудочка от межжелудочковых отверстий до входа в водопровод мозга проходят
гипоталамические борозды, отделяющие таламусы от гипоталамуса. Таламусы соединяются
между собой в средней части III желудочка спайкой – межталамическим
сращением.
б
Рис. 7. Промежуточный и
средний мозг: 1 – мозолистое тело; 2 – полость прозрачной перегородки; 3 –
прозрачная перегородка; 4 – свод (поперечный разрез передних ножек); 5 –
передняя комиссура; 6 – межталамическое сращение; 7 –
задняя комиссура; 8 – бугры четверохолмия (а – верхние, б – нижние);
9 – шишковидная железа; 10 – таламус; 11 – III желудочек; 12 – хвостатое ядро
Промежуточный мозг
включает в себя несколько структур: собственно зрительный бугор – таламус, метаталамус, гипоталамус, субталамус,
эпиталамус, гипофиз.
Таламус (зрительный бугор) – парное крупное образование, является основной частью
промежуточного мозга. Он располагается по обеим сторонам III желудочка и таким
образом формирует его боковые стенки. Таламус включает в себя собственно зрительный
бугор и метаталамус (латеральные и медиальные коленчатые
тела).
Форма таламуса эллипсоидная,
узкая часть направлена назад. Выступающая и более утолщенная задняя часть таламуса
называется подушкой. В передней части зрительный бугор  заканчивается бугорком. Ниже и латеральнее
подушки располагаются продолговато-овальные бугорки: медиальное и латеральное коленчатые
тела.
заканчивается бугорком. Ниже и латеральнее
подушки располагаются продолговато-овальные бугорки: медиальное и латеральное коленчатые
тела.
Медиальная поверхность
таламуса образует боковую стенку III
желудочка, верхняя и латеральная прилегают к внутренней капсуле больших
полушарий, а нижняя граничит с гипоталамусом.
Метаталамус представлен коленчатыми телами, расположенными ниже и
латеральнее подушки. Медиальное коленчатое тело выражено
лучше, лежит под подушкой зрительного бугра и наряду с нижними буграми
четверохолмия является подкорковым центром слуха.
Латеральное коленчатое
тело – небольшое возвышение, лежащее на нижнелатеральной поверхности подушки.
Оно вместе с верхними буграми четверохолмия является подкорковым зрительным центром.
В подушке и коленчатых
телах находятся одноименные ядра. В наружные коленчатые тела входят так
называемые зрительные тракты, которые являются зрительными путями, составленными
уже перекрещенными аксонами ганглиозных клеток сетчатки. Внутренняя структура
таламуса представляет собой ядерные скопления серого вещества, разделенного
белым веществом. В таламусе имеется около 150 ядер. Среди них различают мелкие,
средние и крупные ядра. Их подразделяют на шесть групп: передняя, средней линии,
медиальная, латеральная, задняя и претектальная.
В соответствии с
функциями различают специфические и неспецифические ядра таламуса. Специфические,
в свою очередь, представляют собой переключательные (сенсорные и несенсорные) и
ассоциативные ядра. Аксоны клеток ядер таламуса подходят к определенным участкам
коры.
Переключательные ядра
получают афференты от разных сенсорных систем или от других
отделов мозга, а свои афференты направляют к
определенным проекционным зонам коры. В ассоциативных ядрах заканчиваются афференты от других таламических ядер, а аксоны их клеток
идут к ассоциативным зонам коры. Неспецифические ядра не имеют специфических
афферентных связей с отдельными сенсорными системами, а их афференты
устремляются диффузно ко многим участкам коры.
Переключательные ядра
зрительной и слуховой сенсорных систем – ядра латерального и медиального
коленчатых тел, а соматосенсорной системы – заднее вентральное ядро таламуса. Ассоциативными
ядрами являются латеральные и медиальные ядра подушки.
Неспецифические ядра сосредоточены преимущественно в латеральной, медиальной и
средней группах ядер таламуса.
Таламус связан со
всеми отделами ЦНС. Он является подкорковым центром всех видов общей
чувствительности. Вся информация о состоянии внешней и внутренней среды (за
исключением обонятельной, вкусовой и слуховой) проходит и доходит до сознания
через зрительные бугры. Таламус представляет собой высший центр болевой
чувствительности. Он участвует в переработке сенсорных стимулов, идущих к коре
больших полушарий, а также регулирует цикл бодрствование – сон.
Под таламусом располагается
субталамическая область, которая книзу продолжается в
покрышку ножки мозга. Этот небольшой участок мозгового вещества, отделенный от
таламуса гипоталамической бороздой со стороны III желудочка. В субталамической
области из среднего мозга продолжаются и заканчиваются красные ядра и черная
субстанция. Сбоку от черной субстанции располагается люисово
тело (субталамическое ядро).
Гипоталамус (подбугорная
область) – небольшое образование передненижней части
промежуточного мозга, весом около 4 г. Он образует дно III желудочка и
отделяется от таламусов гипоталамическими бороздами. К гипоталамусу относятся
зрительный перекрест, серый бугор с воронкой и гипофизом, а также сосцевидные
тела.
На основании мозга
между его ножками и сзади от перекреста зрительных нервов (хиазмы) видны
основные структуры гипоталамуса. Серый бугор расположен между сосцевидными телами
и перекрестом зрительных нервов, с боков ограничен зрительными трактами.
Зрительный перекрест имеет вид валика.
Серый бугор соединен с
воронкой, которая переходит вентральнее в ножку
гипофиза и далее в гипофиз. За серым бугром следуют белого цвета парные круглые
образования – сосцевидные тела.
Своей передней частью
гипоталамус прилежит к конечной пластинке мозга, а передненижней – к
зрительному перекресту. Сосцевидные тела граничат с задним продырявленным пространством. Эта часть поверхности мозга называется
продырявленным пространством, потому что пронизана многочисленными сосудами.
Серое вещество
гипоталамуса образует около 30 ядер, которые подразделяются на пять групп: преоптическую, переднюю, среднюю, наружную и заднюю группы.
Преоптическая группа включает медиальное и
латеральное преоптические ядра, а также перивентрикулярное ядро. В переднюю группу входят: супраоптическое, супрахиазматическое
и околожелудочковое ядра. Средняя группа – это
вентромедиальные и дорсомедиальные ядра серого бугра. К наружной группе относят
скопление серого вещества – латеральное гипоталамическое поле. Ядра задней группы
– заднее гипоталамическое, латеральные и медиальные ядра сосцевидных тел:
латеральное мамиллярное ядро меньше медиального. Ядра
гипоталамуса тесно связаны с другими мозговыми структурами. Так, ядра
сосцевидных тел получают большое количество волокон из гиппокампа
(структуры конечного мозга) через свод и из покрышки среднего мозга через мамиллярную ножку.
Эфферентные волокна
сосцевидных тел образуют их главный пучок, или мамиллоталамический, который
направляется к переднему ядру таламуса, а мамиллосегментальный
– к покрышке среднего мозга. Ядра боковых и средних групп гипоталамуса имеют
двусторонние связи с базальными ядрами больших полушарий головного мозга. Ядра
серого бугра получают афференты от обонятельного
мозга, а передняя группа ядер – от свода. Эфференты
ядер гипоталамуса связывают их с таламусом, субталамусом
и другими подкорковыми структурами.
Особо следует выделить
связи передней группы ядер с задней долей гипофиза (нейрогипофизом).
Аксоны клеток этих ядер создают мощный пучок волокон, проходящих в ножке
гипофиза (около 100 тыс. волокон образуют аксоны клеток супраоптического
ядра).
В гипоталамусе различают
три основные группы нейросекреторных клеток: пептидергические;
либерин- и статинергические;
моноаминергические. Однако одни и те же клетки могут синтезировать несколько типов
нейрогормонов.
Паравентрикулярное и супраоптическое ядра
связаны с нейрогипофизом путем прорастания в него аксонов
нервных клеток, образующих эти ядра и формирующих гипоталамо-нейрогипофизарную
систему. В супраоптическом и паравентрикулярном
ядрах синтезируются два пептидных гормона, секретирующихся
из нейрогипофиза. Это вазопрессин и окситоцин.
Гипоталамус является
высшим подкорковым центром интеграции нервных и эндокринных влияний,
вегетативных и эмоциональных компонентов поведенческих реакций и обеспечивает
регуляцию постоянства внутренней среды.
Субталамус. Сзади от гипоталамуса, на задненижнем участке
промежуточного мозга, в межножковой его области над задним продырявленным
пространством находится субталамус. В этой части
промежуточного мозга у человека располагается субталамическое
ядро Люиса.
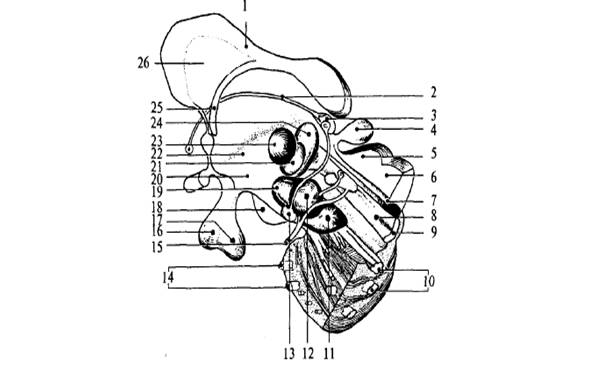
Рис.
8. Схема основных ядер промежуточного и среднего мозга: 1 – мозолистое тело; 2 – мозговая полоска зрительного
бугра; 3 – уздечка; 4 – эпифиз; 5 – верхний бугорок четверохолмия; 6 – нижний
бугорок четверохолмия; 7 – Сильвиев водопровод; 8 –
медиальный продольный пучок; 9 – медиальная петля; 10 – пирамидный тракт; 11 –
черная субстанция среднего мозга; 12 – красное ядро; 13 – межножковое ядро; 14
– поперечные волокна моста; 15 – глазодвигательный нерв; 16 – аденогипофиз; 17 – нейрогипофиз;
18 – сосцевидное тело; 19 – субталамическое ядро; 20
– гипоталамус; 21 – вентропостеромедиальное ядро
таламуса; 22 – гипоталамическая борозда; 23 – центральное медиальное ядро
таламуса; 24 – вентропостеролатеральное ядро
таламуса; 25 – свод; 26 – прозрачная перегородка
Эпиталамус. По границам эпителиальной пластинки и боковым стенкам
III желудочка протянулись мозговые полоски, расширяющиеся сзади в треугольники
поводка. B этих треугольниках серое вещество формирует ядра поводка. Треугольники
поводка направляются к середине дорсальной поверхности среднего мозга и
переходят в поводки. Соединяясь, поводки образуют спайку, к задней поверхности
которой прикреплено шишковидное тело, или эпифиз.
Мозговые полоски,
треугольники, поводки и эпифиз образуют эпиталамус.
Эпифиз является железой внутренней секреции, контролирующей течение
циркадианных ритмов и тормозящей активность половых желез до наступления
периода полового созревания.
Гипофиз, или
нижний мозговой придаток, располагается на вентральной поверхности мозга в
основании черепа в ямке турецкого седла. После рождения по мере роста и
развития ребенка масса гипофиза увеличивается и к 14 годам достигает массы гипофиза
взрослого человека:
Гипоталамус и гипофиз
составляют единую функциональную систему, обеспечивающую совместную регуляцию
функций нервным и гуморальным путем. По своему строению и эмбриогенезу гипофиз
не однороден. В гипофизе различают две главные части: нейрогипофиз
и аденогипофиз, имеющие различное эмбриональное происхождение
и строение.
Нейрогипофиз представляет
собой производное дна воронки
промежуточного мозга. Он
находится в тесной морфологической и функциональной связи с гипоталамусом, в
нем заканчиваются волокна гипоталамо-гипофизарного тракта, идущего от супраоптического и паравентрикулярного
ядер гипоталамуса.
Аденогипофиз (передняя
доля) развивается из эпителиального выпячивания (кармана Ратке)
крыши кишечной трубки.
Передняя доля гипофиза
имеет тесную сосудистую связь с гипоталамусом. Здесь артерии разветвляются на
капилляры, образуя плотное сплетение в форме мантии на поверхности срединного
возвышения. Капиллярные ветви этого сплетения образуют вены, достигающие передней
доли гипофиза, здесь вены вновь распадаются на капилляры, пронизывающие всю
долю. Вся эта сложная система кровеносных сосудов носит название портальной. По
ней в аденогипофиз из гипоталамуса поступают пептидные
гормоны (либерины и статины),
регулирующие синтез и секрецию гормонов аденогипофиза.
Нейрогипофиз имеет собственную систему кровоснабжения,
не зависящую от портальной системы.
В аденогипофизе
секретируется два типа гормонов – эффекторные,
т. е. реализующие свои свойства непосредственно в организме, и тропные – оказывающие регулирующее влияние на
периферические железы внутренней секреции. Всего в аденогипофизе
синтезируется шесть гормонов – гормон роста, пролактин, тиреотропин,
адренокортикотропный гормон (АКТГ), фолликулостимулирующий гормон, лютеинизирующий гормон. Фолликулостимулирующий и лютенизирующий гормоны объединяются в группу гонадотропных
гормонов.
За последние годы было
установлено, что практически все биологически активные вещества, секретируемые
нейронами гипоталамо-гипофизарной системы, имеют пептидную природу.
Филогенез. В филогенезе структуры промежуточного мозга появляются
и развиваются на разных его этапах. У низших позвоночных животных уже есть
некоторые эпиталамические, метаталамические
и гипоталамические структуры. У круглоротых таламус уже достаточно выражен и
разделяется на дорсальную и вентральную части, включающие в себя несколько
ядер. Гипоталамус является также выраженной структурой. У костистых рыб
происходят усложнение в организации таламуса и дифференциация ряда ядерных
групп. У амфибий и рептилий развиваются зрительные бугры. Они становятся
специальными структурами и связывают средний мозг с конечным. У рептилий
промежуточный мозг выполняет вместе с некоторыми структурами конечного мозга
высшие интегративные функции. Дорсальный таламус у них отделяется от вентрального,
на этом этапе выделяются четкие ядра с характерным строением и связями. В дорсальном
таламусе их насчитывается 9, в вентральном – 7. В переднем отделе гипоталамуса
у рептилий появляются зачатки супраоптического и паравентрикулярного ядер. У птиц дифференциация ядер гипоталамуса
хорошо выражена, тогда как таламус усложняется незначительно.
Особенно бурно идет
развитие дорсального таламуса у млекопитающих, так как он выполняет функцию
основного коллектора сенсорных путей к коре больших полушарий. У млекопитающих
наряду с развитием релейных (переключательных) ядер таламуса развиваются
ассоциативные ядра. Происходит усиленное развитие таламуса и формируется таламонеокортикальная система интеграции.
Онтогенез. В
онтогенезе структуры промежуточного мозга формируются из стенок второго мозгового
пузыря пятипузырной стадии развития головного мозга. Это
происходит следующим образом: глазные пузырьки превращаются в глазные бокалы и
глазные стебельки. Позже из них строятся сетчатка глаза и зрительные проводящие
пути. Из боковых стенок второго пузыря образуются структуры дорсального и
вентрального таламусов. Верхней стенкой являются эпиталамические
структуры, нижней – гипоталамус. Из верхней части задней стенки развиваются забугорные
структуры. B этой части мозговой трубки все структуры образуются во время
онтогенеза из крыловидной пластинки (пограничная борозда и основная пластинка
заканчиваются на уровне среднемозгового пузыря). Поэтому здесь не возникают ни
моторные (двигательные), ни вегетативные ядра, а следовательно, нет и
соответствующих черепно-мозговых нервов. Все ядра промежуточного мозга являются
либо сенсорными (переключательными), либо интегративными (ассоциативными).
Имеются здесь и неспецифические
ядра. Гипоталамус отделяется от соседних участков мозга на 2-м месяце
внутриутробной жизни. Вслед за этим начинается формирование шести гипоталамических
ядер, которые представляют собой скопление нейронов с определенными функциями.
Дифференцировка входящих в них клеток продолжается до 6-го месяца
внутриутробной жизни, а завершается еще позднее. В четырех из шести ядер
вырабатываются гормоны, которые по системе сосудов направляются в аденогипофиз.
Гипоталамо-аденогипофизарную систему составляют супрахиазматическое
ядро, вентромедиальное, дорсомедиальное и аркуатное
ядра. Сосудистая система появляется на 14-й неделе в виде первых капиллярных
петель, и ее формирование завершается к моменту рождения. Синтезируемые в этих
ядрах регуляторные пептиды обнаруживаются в аденогипофизе
уже на 10-й неделе развития плода. Однако по некоторым наблюдениям в первые три
месяца внутриутробной жизни, а возможно, до первой половины беременности,
гипофиз не подчиняется контролю гипоталамуса. Это объясняется незрелостью
нейросекреторных клеток и недостаточным развитием портальной системы сосудов.
Вазопрессин появляется
в гипофизе плода на 15–17-й, а окситоцин – на 18–19-й неделях внутриутробного
развития. К 6-му месяцу внутриутробной жизни содержание их значительно возрастает.
Уже в этот период они принимают участие в регуляции жизнедеятельности плода. Установление
гипоталамического контроля над эндокринными железами происходит к концу внутриутробного
развития.
Тема 6. Базальные
ганглии (подкорковые ядра).
Лимбическая система
В основании больших
полушарий (нижняя стенка боковых желудочков) расположены ядра серого вещества
– базальные ганглии. Они составляют
примерно 3 % от объема полушарий. Подкорковые (базальные, центральные) ядра
представляют собой образования, филогенетически более ранние, чем кора больших
полушарий. Помимо коры, образующей поверхностные слои конечного мозга, серое
вещество в каждом из полушарий залегает в виде отдельных скоплений – ядер, или
узлов, которые располагаются в толще белого вещества, находясь ближе к основанию
полушарий большого мозга. Название базальные, или
подкорковые, ядра объясняется их анатомическим положением. Они представлены
хвостатым, чечевицеобразным ядрами, оградой и миндалиной.
Все базальные ганглии
функционально объединены в две системы. Первая группа ядер представляет собой стриопаллидарную
систему, включающую хвостатое
ядро, скорлупу и бледный шар. Скорлупа и хвостатое ядро имеют
слоистую структуру и поэтому их общее название – полосатое тело. Такое название – полосатое тело – связано с
тем, что на срезах наблюдается чередование белого и серого веществ.
Бледный шар не имеет
слоистости и выглядит светлее стриатума. Скорлупа и бледный
шар объединены в чечевицеобразное ядро. По своей форме чечевицеобразное ядро напоминает трехгранную пирамиду,
обращенную свободным образованием наружу. Ядро отчетливо делится прослойками
белого вещества на темноокрашенную латеральную часть – скорлупу и боле светлую
часть – бледный шар, состоящую из двух фрагментов: наружного и внутреннего.
Скорлупа образует
наружный слой чечевицеобразного ядра, а бледный шар – внутренние его части.
Скорлупа по генетическим, структурным и функциональным признакам близка к хвостатому
ядру. Анатомически хвостатое ядро тесно связано с боковым желудочком. Хвостатое
ядро вытянуто в сагиттальной плоскости и сильно изогнуто. Его передняя
утолщенная часть – головка – находится впереди таламуса. Постепенно, суживаясь,
сзади оно переходит в длинный хвост. Расположенная впереди и медиально
расширенная его часть – головка хвостатого ядра – образует боковую стенку
переднего рога желудочка, тело ядра – нижнюю стенку центральной части желудочка,
а тонкий хвост – верхнюю стенку нижнего рога. Следуя форме бокового желудочка,
хвостатое ядро охватывает дугой чечевицеобразное ядро. Друг от друга хвостатое и чечевицеобразное
ядра отделены прослойкой белого вещества – частью внутренней капсулы.
Другая часть внутренней капсулы отделяет чечевицеобразное ядро от нижележащего
таламуса.
Таким образом,
строение дна бокового желудочка (представляющего собой стриопаллидарную
систему) схематично можно представить себе так: стенку самого желудочка
образует слоистое хвостатое ядро, затем ниже идет прослойка белого вещества – внутренняя
капсула, под ней слоистая скорлупа, еще ниже бледный шар и опять слой
внутренней капсулы, лежащий на ядерной структуре промежуточного мозга – таламусе.
Стриопаллидарная система получает афферентные волокна от
неспецифических медиальных таламических ядер, лобных отделов коры больших
полушарий, коры мозжечка и черной субстанции среднего мозга. Основная масса
эфферентных волокон стриатума радиальными пучками
сходится к бледному шару.
Таким образом, бледный
шар является выходной структурой стриопаллидарной
системы. Эфферентные волокна бледного шара идут к передним ядрам таламуса,
которые связаны с фронтальной и теменной корой больших полушарий. Часть эфферентных
волокон, не переключающихся в ядре бледного шара, идет к черной субстанции и красному
ядру среднего мозга.
Стриопаллидум совместно со своими проводящими путями, входит в экстрапирамидную систему, оказывающую тоническое влияние на
моторную деятельность. Эта система контроля над движениями называется
экстрапирамидной потому, что переключается на пути к спинному мозгу, минуя
пирамиды продолговатого мозга.
Стриопаллидарная система является высшим центром непроизвольных и
автоматизированных движений, снижает мышечный тонус, тормозит движения,
осуществляемые двигательной корой. Латеральнее стриопаллидарной системы базальных ганглиев расположена
тонкая пластинка серого вещества ограда, расположенная вблизи скорлупы. Она
ограничена со всех сторон волокнами белого вещества – наружной капсулой. Слоем
белого вещества ограда отделена от коры островка.
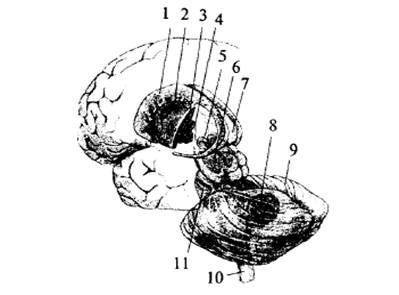
Рис.
9. Базальные ядра, их положение и структуры ствола мозга: 1 – хвостатое ядро; 2
– скорлупа; 3 – бледный шар; 4 – внутренняя капсула; 5 – субталамическое
ядро; 6 – черная субстанция; 7 – таламус; 8 – подкорковые ядра мозжечка; 9 –
мозжечок; 10 – спинной мозг; 11 – мост
В процессе филогенеза
полосатое тело расположилось над средним мозгом. Получая импульсы от зрительных
бугров, эти ядра принимают участие в выполнении сложных автоматических движений
(ходьба, бег, лазание и т. д.). В ядрах полосатого тела замыкаются дуги сложных
безусловных рефлексов. Полосатое тело играет важную роль в запоминании двигательных
программ. Повреждение или нарушение этой структуры приводит к ухудшению памяти
и обучения. Кроме того, полосатое тело способствует повышению степени
проявления агрессивных реакций.
Подкорковые ядра
связаны не только с двигательными реакциями, но и вегетативными функциями. Они
являются высшими центрами автономной нервной системы. При повреждении
нисходящих путей, связанных с базальными ядрами, отмечается неподвижность тела,
отказ от еды и питья, отсутствуют реакции на раздражения, поступающие из внешнего
мира.
Остальные базальные
ядра входят в лимбическую систему мозга.
Впереди от конца нижнего рога бокового желудочка в белом веществе височной доли
больших полушарий расположена плотная группа ядер – миндалевидное тело. Амигдала (то же, что и миндалина) – анатомический термин,
обозначающий субкортикальное базальное ядро, которое располагается в толще
височной доли (по одному в каждом полушарии), впереди от верхушки нижнего рога. Амигдалу включают в
состав лимбической системы, она имеет тесные связи с
другими частями этой системы. Она также получает сигналы от всех сенсорных
систем. С функционированием амигдалы связаны:
эмоциональное поведение (страх, оборонительное и агрессивное поведение),
мотивация и сохранение части прошлого опыта. Миндалевидное тело принимает также
импульсы от разнообразных афферентных систем, участвует в тонком и точном
проявлений эмоциональных реакций.
В пределах прозрачной
перегородки лежит ядро перегородки. Кроме перечисленных базальных ядер,
в лимбическую систему входит совокупность
функционально связанных между собой образований древней коры (гиппокамп, грушевидная доля, энторинальная
область, периамигдалоидная кора), а также старой коры
(поясная извилина, пресубикулюм) и подкорковых
структур (миндалевидный комплекс, область перегородки, ряд ядер, таламуса и
гипоталамуса, а также лимбическая зона среднего
мозга). Эта система образует нечто вроде кольца, состоящего из таких пучков
нервных волокон, как, например, свод и поясной пучок, соответствующий поясной
извилине. Эти пучки соединяют некоторые ядра передней части гипоталамуса с гиппокампом и расположенным в височной доле миндалевидным
телом.
Таким образом, лимбическая система тесно связана с гипоталамусом, вместе с
которым она выполняет важные функции, касающиеся мотивации и эмоций, которые
она контролирует. Лимбическая система участвует в
управлении эмоциональным и инстинктивным поведением (пищевым, половым,
оборонительным). Эта система является важным звеном регуляторной системы,
модулирующим уровень активации коры больших полушарий. Структуры ее (гиппокамп, миндалевидный комплекс и др.) играют важнейшую
роль в потребностно-эмоциональных и мнестических (приемы, связанные с увеличением объема памяти,
облегчение запоминания) процессах. С функцией гиппокампа
связывают перенос кратковременной памяти в долговременную.
Свои нисходящие пути лимбическая система направляет к ретикулярной формации
ствола мозга и к гипоталамусу. Через гипоталамо-гипофизарную систему лимбическая система осуществляет контроль над гуморальной
системой. Для лимбической системы характерна особая
чувствительность и особая роль в ее функционировании гормонов, синтезируемых в
гипоталамусе и секретируемых гипофизом, – окситоцина и вазопрессина.
Основной функцией лимбической системы является осуществление эмоционально
мотивационного поведения. Она организует и обеспечивает протекание
вегетативных, соматических и психических процессов при
эмоционально-мотивационной деятельности, а также осуществляет восприятие и
хранение эмоционально значимой информации, выбор и реализацию адаптивных форм
эмоционального поведения.
Вместе с тем каждая
структура, входящая в лимбическую систему, вносит
свой вклад в единый механизм, имея свои функциональные особенности. Так,
функции гиппокампа связаны с памятью, обучением, формированием
новых программ поведения при изменении условий. Передняя лимбическая
кора обеспечивает эмоциональную выразительность речи, перегородка принимает
участие в переобучении, снижает агрессивность и страх. Мамиллярные
тела играют большую роль в выработке пространственных навыков, миндалевидный
комплекс отвечает за пищевое и оборонительное поведение.
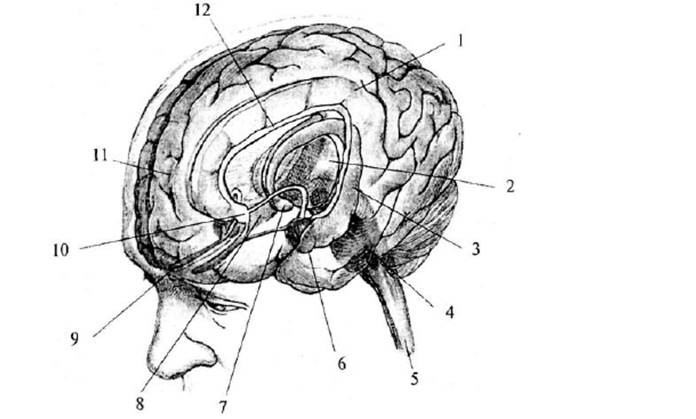
Рис.
10. Лимбическая система: 1 – поясная извилина; 2 – таламус;
3 – левый гиппокамп; 4 – ретикулярная формация; 5 – спинной
мозг; 6 – миндалина; 7 – мамиллярное тело; 8 – правый
гиппокамп; 9 – обонятельная луковица; 10 – перегородка;
11 – лобная доля; 12 – свод
Лимбическая система организует и обеспечивает протекание
вегетативных, соматических и психических процессов при эмоционально-мотивационной
деятельности, а также осуществляет восприятие и хранение эмоционально значимой
информации, выбор и реализацию адаптивных форм эмоционального поведения. Вместе
с тем каждая структура, входящая в лимбическую систему,
вносит свой вклад в единый механизм, имея свои функциональные особенности. Так,
функции гиппокампа связаны с памятью, обучением,
формированием новых программ поведения при изменении условий. Передняя лимбическая кора обеспечивает эмоциональную выразительность
речи, перегородка принимает участие в переобучении, снижает агрессивность и
страх. Мамиллярные тела играют большую роль в
выработке пространственных навыков, миндалевидный комплекс отвечает за пищевое
и оборонительное поведение.
Лимбическая система и особенно входящий в нее гиппокамп
играют важную роль и в процессах памяти.
Кроме того, она участвует в регуляции агрессивного поведения, которое можно
вызывать стимуляцией миндалевидного ядра и подавлять раздражением области
перегородки расположенной впереди свода. В различных отделах лимбической системы, окружающих гиппокамп,
были также обнаружены центры удовольствия и боли. Входящие в лимбическую систему структуры действуют сообща, т. е.
возбуждение, возникающее в одной структуре, тут же охватывает другие.
Половое влечение, голод, жажда – эти наиглавнейшие побудительные
причины деятельности всех живых существ связаны, прежде всего, именно с лимбической системой. Так в гипоталамусе располагаются
группы клеток, реагирующих на изменения уровня питательных веществ и воды в
крови. При низком содержании «еды» в крови эти клетки тут же передают «тревожные»
сигналы в высшие отделы коры головного мозга, в результате чего возникают
чувства голода и жажды, которые и заставляют организм активно заняться поиском
пропитания.
При поражении лимбического
отдела мозга часто возникают двигательные и психические реакции, которые могут
быть абсолютно противоположны: или беспокойство, настороженность, агрессия,
стремление бежать, или, наоборот, спокойствие, пассивность, умиротворенность. Это
объясняется тем, что лимбическая система участвует в
приспособительных реакциях, сложившихся у наших далеких предков на ранних
стадиях эволюции, еще тогда, когда в критических и опасных ситуациях могло быть
лишь два варианта спасения: активный – убегать или нападать и пассивный – замаскироваться,
спрятаться, затихнуть и замереть. Именно так до сих пор поступают многие
низкоорганизованные животные.
Следовательно, главнейшее место в приспособительной деятельности
принадлежит эмоциям, биологический смысл и биологическое предназначение которых
и заключается в быстрой оценке текущих потребностей организма и стимуляции
соответствующего ответа на действие того или иного раздражителя.
Именно в лимбической системе,
в основном в гипоталамусе, формируются эмоции. Изменения в лимбических
структурах, возникающие при определенных стрессовых состояниях, неврозах,
иногда в результате опухоли или нарушения мозгового кровообращения или инфекционного
заболевания, могут повлечь за собой и нарушение эмоционального равновесия. Поэтому
при наличии различных патологий преобладать будут отрицательные эмоции – страх,
напряжение, тоска, беспричинная тревога.
Конечно, возможны и прямо противоположные реакции – чрезмерно
повышенное настроение, двигательная активность, переоценка своих возможностей,
но на этом уже скажется поражение миндалевидного комплекса. Имеются данные о
том, что развитие таких заболеваний, как ишемическая болезнь сердца, гипертоническая
и язвенная болезни, во многом связано с отрицательными эмоциями. Нормализуя
эмоциональные реакции человека, можно избавить его от многих болезней. На этом
принципе и построен эффект психотропных средств, которые прежде всего
воздействуют на лимбическую систему, а уже через нее
– на функции сердца, сосудов, органов пищеварения.
Лимбическая система, в основном гиппокамп,
принимает активное участие в сложнейших процессах, лежащих в основе памяти. Однако
гиппокамп не является длительным хранилищем
поступающей в мозг информации, т. к. эту роль выполняет кора больших полушарий,
но зато из-за особенностей анатомического строения вся лимбическая
система как будто создана для кратковременного хранения информации. Благодаря
переплетению пучков аксонов, соединяющих различные образования лимбической системы, в ней формируется ряд больших и малых
замкнутых кругов, приспособленных для повторного циркулирования
нервных импульсов и сохранения возбуждения в течение определенного времени.
Случаи повреждения гиппокампа
или хирургического его удаления подтверждают, что эта структура играет решающую
роль при запоминании новых событий и хранении их в долговременной памяти, но не
для воспроизведения старых воспоминаний. Например, после удаления гиппокампа больной без труда узнает старых друзей, помнит
свое прошлое, может читать и пользоваться ранее приобретенными навыками, но
зато он не помнит о том, что происходило
в течение примерно года до операции. А вот события или людей, которых он встретил
после операции, не будет помнить вообще. Такой пациент не сможет узнать нового
человека, с которым он провел много часов ранее в этот же день. Он будет неделю
за неделей собирать одну и ту же головоломку и никогда не вспомнит, что уже собирал
ее раньше, будет снова и снова читать ту же газету, не помня ее содержания.
При поражении гиппокампа
алкоголем, у человека также нарушается память на недавние события. Как
показывают наблюдения врачей, алкоголики, находящиеся на лечении в больнице,
затрудняются ответить на вопросы о том, обедали они сегодня или нет, когда
принимали лекарство, работали ли в мастерской. Однако давние события своей
жизни они помнят хорошо.
Тема 7. Конечный мозг
Конечный мозг
– самый большой отдел головного мозга. Он состоит из двух полушарий, которые разделены продольной мозговой щелью. Полушария
сильно разрастаются и покрывают собой все остальные отделы головного мозга.
Длина полушарий
Подпаутинное пространство составляет функционально единое целое с
венозной системой мозга и его желудочками. Каждое полушарие имеет свою
щелевидную полость – боковой желудочек.
Условно желудочек левого полушария считается I, а правого – II. Вслед за ростом
полушарий боковые желудочки разрастаются и принимают своеобразную форму.
Кроме центральной
части, каждый желудочек имеет распространяющийся в лобную долю передний рог, идущий
в глубине затылочной доли задний рог и отходящий от него вниз и латерально нижний рог, являющийся полостью височной доли.
Боковые желудочки сообщаются с полостью III желудочка и имеют в своих стенках
сосудистое сплетение. Медиальной стенкой каждого переднего рога является тонкая
прозрачная пластинка. Правая и левая пластинки образуют между передними рогами
общую прозрачную перегородку.
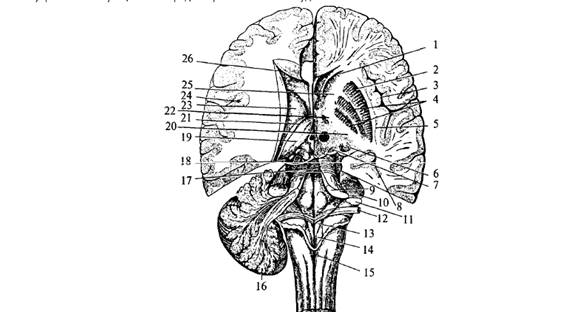
Рис.
11. Большие полушария мозга и базальные ядра: 1 – головка хвостатого ядра; 2 – скорлупа; 3 – кора мозгового островка;
4 – бледный шар; 5 – ограда; 6 – хвост хвостатого ядра; 7 – ядра медиального
коленчатого тела; 8 – нижний рог бокового желудочка; 9 – верхние ножки
мозжечка; 10 – средние ножки мозжечка; 11 – нижние ножки мозжечка; 12 – мозговые
полоски (волокна проводящих путей слуховой системы); 13 – треугольник
подъязычного нерва; 14 – треугольник блуждающего нерва; 15 – задвижка; 16 – часть мозжечка; 17 – верхний мозговой парус;
18 – блоковый нерв; 19 – таламус; 20 – задняя ножка внутренней капсулы;
21 – конечная пластинка; 22 – колено внутренней капсулы; 23 – хвостатое ядро;
24 – кора мозгового островка; 25 – передняя ножка внутренней капсулы; 26 – передний
рог бокового желудочка
Расположение белого и
серого веществ в полушариях сходно с таковым в мозжечке: в глубине белого
вещества расположены ядра серого вещества, а вся поверхность полушарий покрыта
серым веществом (плащом). Толщина коры в различных участках неодинакова и
колеблется в пределах от 1,5 до
Размеры, форма и
взаиморасположение нейронов, составляющих
новую кору, отличаются в различных участках новой коры и служат основой для
определения ее нейроцитоархитектоники.
Под нейроцитоархитектоникой понимают особенности микроскопического
строения коры. Различают цитоархитектонику и миелоархитектонику. Цитоархитектоника
– совокупность особенностей клеточного состава, миелоархитектоника
– совокупность особенностей волокнистого строения. Волокнистое строение в
основном соответствует клеточному составу.
Начало изучения
архитектоники коры головного мозга относится к концу XVIII века, когда в
По происхождению и
структуре кора больших полушарий неоднородна. Большую часть коры у человека
занимает новая кора – неокортекс,
филогенетически наиболее молодая корковая формация. Филогенетически более
ранние корковые структуры – древняя кора и старая кора – занимают
небольшую часть поверхности полушарий.
Закладка новой коры
происходит в латеральных частях плаща. Новая кора интенсивно развивается и
оттесняет древнюю кору на основание
полушарий, где она сохраняется в виде узкой полоски обонятельной коры и
занимает 0,6 % поверхности коры на вентральной поверхности полушарий, а старая
кора отодвигается на медиальные поверхности полушарий, занимает 2,2 % поверхности
коры и представлена гиппокампом и зубчатой извилиной.
По происхождению и
клеточному строению новая кора отличается от древней и старой коры. Однако резких
цитоархитектонических границ между ними нет. Переход от одной корковой формации
к другой в клеточном строении происходит постепенно. Кора переходного типа
называется межуточной корой, она занимает 1,3 % общей площади коры.
Таким образом, большую часть поверхности коры (95,6 %) занимает новая кора.
Древняя и старая кора. Для
древней коры характерно отсутствие послойного строения. В ней преобладают
крупные нейроны, сгруппированные в клеточные островки. Старая кора имеет три
клеточных слоя. Ключевой структурой старой коры является гиппокамп.
Гиппокамп, или аммонов рог,
расположен медиобазально в глубине височных долей. Он
имеет своеобразную изогнутую форму (гиппокамп в
переводе – морской конек) и почти на всем своем протяжении образует впячивание в полость нижнего рога бокового желудочка.
Гиппокамп является собственно складкой (извилиной) старой коры.
С ней сращена и заворачивается над ней зубчатая извилина. Являясь частью старой
коры, гиппокамп имеет слоистую структуру. К зубчатой
извилине примыкает слой конечных ветвлений апикальных дендритов.
Серое вещество
расположено на поверхности полушарий и покрывает новую кору, занимающую у
человека 96 % всей коры. Все области новой коры построены по единому принципу.
Исходным типом является шестислойная кора. Слои представлены следующим образом:
![]() Рис. 12. Схема строения новой коры: I – молекулярный слой; II – наружный зернистый слой;
III – пирамидный слой; IV – внутренний зернистый слой; V – ганглионарный
слой; VI – полиморфный слой
Рис. 12. Схема строения новой коры: I – молекулярный слой; II – наружный зернистый слой;
III – пирамидный слой; IV – внутренний зернистый слой; V – ганглионарный
слой; VI – полиморфный слой
I слой – молекулярный. Этот слой является самым
поверхностным, имеющим толщину около
II слой – наружный зернистый. Толщина
наружного зернистого слоя – 0,10 мм. Он состоит из мелких пирамидных и звездчатых
нейронов. Аксоны этих нейронов оканчиваются на нейронах III, V и VI слоев.
III слой – пирамидный, он имеет толщину около
IV слой – внутренний
зернистый. Он состоит
из звездчатых клеток, имеющих короткие отростки, и малых пирамид. Дендриты
клеток IV слоя уходят в молекулярный слой коры, а их коллатерали ветвятся в
своем слое. Аксоны клеток IV слоя могут подниматься в вышележащие слои или
уходить в белое вещество как ассоциативные волокна. Толщина IV слоя от 0,12 до
V слой – ганглионарный – слой
больших пирамидных нейронов. Самые крупные клетки коры расположены именно в
этом слое (гигантские пирамиды Беца передней
центральной извилины). Их апикальные дендриты достигают молекулярного слоя, а
базальные дендриты распределяются в своем слое. Аксоны клеток V слоя покидают
кору и являются ассоциативными, комиссуральными или
проекционными волокнами. Толщина V слоя достигает
VI слой коры – полиморфный. Содержит клетки разнообразной формы и размера, имеет
толщину от 0,1 до
Однако не вся кора
построена таким образом. На медиальной и нижней поверхности полушарий сохранились
участки древней и старой коры, имеющие трехслойное строение.
Все клетки коры
являются вставочными нейронами рефлекторных дуг. По своему строению и характеру
участия в деятельности коры среди них различают нейроны с длинными и короткими
нейритами. Клетки с длинными аксонами представлены пирамидными и
веретеновидными нейронами. Они встречаются в V и VII слоях. Длинный нисходящий аксон их отдает многочисленные
коллатерали на всем своем пути следования и, покидая кору, продолжается в виде
белого вещества как нисходящее проекционное волокно.
Проекционное волокно
оканчивается в следующих структурах: базальных ганглиях, двигательных ядрах ствола
мозга или на мотонейронах спинного мозга. Восходящий
дендрит пирамидных клеток доходит до первого слоя и там формирует густое ветвление,
в результате чего эти нейроны собирают информацию со всех слоев коры. В верхних
слоях длинные нейриты имеют нейроны III слоя.
Аксоны этих клеток входят в состав белого вещества преимущественно в качестве
ассоциативных и комиссуральных волокон.
Нейроны с короткими
нейритами характеризуются тем, что их аксоны не выходят за пределы коры. По
форме они звездчатые, кустовидные и паукообразные. Они
встречаются во всех слоях коры. В коре больших полушарий человека этих нейронов
гораздо больше, чем у других млекопитающих. В IV слое они являются главными клеточными элементами. Их
роль заключается в восприятии афферентных импульсов и затем распределении их на
пирамидные клетки III и V слоев. Кроме того, звездчатые клетки осуществляют
круговую циркуляцию импульсов в коре: восприняв нервный импульс, они могут
долгое время находиться в состоянии скрытой активности, т. е. не проявляя
внешне видимой реакции. Это служит одной из форм материального субстрата
памяти, анатомо-функциональной предпосылки для динамической фиксации следов
возбуждения, удержания и эффективного использования информации, запасаемой
человеком на протяжении всей его жизни.
Сложность и
совершенство коры больших полушарий человека по сравнению с животными зависит
главным образом от увеличения количества
нейронов с короткими нейритами. Значительное же увеличение площади коры
большого мозга связано с ростом и развитием ассоциативных волокон.
![]()
Рис.
13. Медиальная поверхность левого полушария: 1 – предцентральная
извилина (двигательные зоны); 2 – поясная извилина (часть лимбической
системы), отвечает за висцеральную чувствительность; 3 – мозолистое тело
(основная комиссура); 4 – свод; 5 – лобная доля; 6 – обонятельные нервы,
обонятельная луковица и обонятельный путь; 7 – височная доля; 8 – гиппокамп (часть лимбической
системы); 9 – первичное проекционное зрительное поле (17 поле); 10 – вторичное
проекционное зрительное поле (18 поле); 11 – затылочная доля; 12 – теменная
доля; 13 – задняя центральная извилина (соматосенсорные зоны)
По ширине слоев,
форме, величине и плотности расположения нейронов кору делят на области и поля.
Области коры появляются в процессе развития раньше, чем поля, выделяющиеся в
ходе структурной дифференцировки, и характеризуются примерно одинаковыми
признаками.
Структуры конечного
мозга, лежащие над стриатумом (крыша, латеральные и
медиальные стенки боковых желудочков), носят название плаща. Именно
плащ, значительно разрастаясь, образуя складки своей поверхности, закрывает
собой почти все отделы головного мозга. Поверхностный слой плаща, состоящий из
серого вещества, называется корой больших полушарий. Площадь поверхности
обоих полушарий около 1650 см2.
Каждое полушарие имеет
три поверхности – верхнелатеральную,
медиальную и нижнюю. Крупными бороздами каждое полушарие делится на доли. Центральная, или Роландова борозда, расположена в верхней
части латеральной поверхности полушария и отделяет лобную долю от теменной. Латеральная, или Сильвиева
борозда, идет также по латеральной поверхности полушария и отделяет височную
долю от лобной и теменной. Теменно-затылочная борозда отделяет теменную и
затылочную доли по медиальной поверхности полушарий. В глубине Сильвиевой борозды лежит островковая доля, закрытая со всех
сторон «наползшими» на нее участками коры. Кроме того, часто выделяют еще одну
долю, которая расположена в глубине медиальной поверхности полушария и
дугообразно охватывает промежуточный мозг. Это лимбическая
доля. Более мелкие борозды делят доли на извилины. Часть этих борозд постоянна
(наблюдается у всех индивидов), другая индивидуальна (наблюдается не у всех и
не всегда). 2/3 поверхности коры образуют боковые стенки борозд и только 1/3
находится на поверхности извилин.
Клетки одного слоя
коры выполняют сходную функцию в обработке информации. I и IV слои являются
местом ветвления ассоциативных и комиссуральных
волокон, т. е. получают информацию от других корковых структур. III и IV слои
являются входными, афферентными для проекционных полей, так как в этих слоях
заканчиваются таламические волокна. V слой клеток выполняет эфферентную
функцию, его аксоны несут информацию к нижележащим структурам мозга. VI слой
также является выходным и ассоциативным, но его аксоны не покидают кору.
Основным принципом
функциональной организации коры является объединение нейронов в колонки.
Колонка расположена перпендикулярно поверхности коры и охватывает все ее слои
от поверхности к белому веществу. Связи между клетками одной колонки
осуществляются по вертикали вдоль оси колонки. Боковые отростки клеток имеют
небольшую длину. Связь между колонками соседних зон осуществляется через
волокна, уходящие вглубь, а затем входящие в другую зону, т. е. короткие ассоциативные
волокна. Функциональная организация коры в виде колонок обнаружена в
соматосенсорной, зрительной, моторной и ассоциативной коре.
Отдельные зоны коры
имеют принципиально одинаковое клеточное строение, однако есть и отличия,
особенно в структуре III, IV и V слоев, которые могут распадаться на несколько подслоев.
Кроме этого, существенными цитоархитектоническими признаками являются плотность
расположения и размеры клеток, наличие специфических типов нейронов,
расположение и направление хода миелиновых волокон. Цитоархитектонические
особенности позволили разделить всю поверхность коры на 11 цитоархитектонических
областей, включающих в себя 52 поля (по Бродману).
Каждое цитоархитектоническое поле обозначено на картах мозга номером, который
присваивался ему в порядке описания. Следует отметить, что между
цитоархитектоническими полями не существует резких границ, клеточные слои плавно
меняют свою структуру при переходе от одного поля к другому. Каждое поле коры
выполняет определенную функцию. Часть полей коры являются сенсорными.
В первичных сенсорных
полях заканчиваются проекционные афферентные волокна. Из первичных сенсорных полей
информация по коротким ассоциативным волокнам передается во вторичные проекционные
поля, расположенные рядом с ними. Так, поля 1 и 3, занимающие медиальную и латеральную
поверхность задней центральной извилины, являются первичными проекционными полями
кожной чувствительности противоположной половины поверхности тела. Расположенные
рядом друг с другом участки кожи также рядом друг с другом проецируются на корковой
поверхности. Такая организация проекций называется топической.
В медиальной части представлены
нижние конечности, а наиболее низко на латеральной части извилины расположены проекции
рецепторных полей кожной поверхности головы. При этом участки поверхности тела,
богато снабженные рецепторами (пальцы, губы, язык), проецируются на большую
площадь коры, чем участки, имеющие меньшее количество рецепторов (бедро, спина,
плечо). Поле 2, расположенное в нижнелатеральной части той же извилины,
является вторичным проекционным полем кожной чувствительности. Поля 17–19,
расположенные в затылочной доле, являются зрительным центром коры, 17-е поле, занимающее
сам затылочный полюс, является первичным. Прилежащие к нему 18 и 19-е поля выполняют
функцию вторичных ассоциативных полей и получают входы от 17-го поля. В височных
долях расположены слуховые проекционные поля 41, 42. Рядом с ними на границе височной,
затылочной и теменной долей расположены 37, 39 и 40-е, характерные только для коры
головного мозга человека. У большей части людей в этих полях левого полушария расположен
центр речи, отвечающий за восприятие устной и письменной речи. Поле 43, занимающее
нижнюю часть задней центральной извилины, получает вкусовые афференты.
Структуры обонятельной
чувствительности посылают свои сигналы в кору больших полушарий без
переключений в других отделах ЦНС. Обонятельные луковицы расположены под нижней
поверхностью лобных долей. От них начинается обонятельный тракт, который является
первой парой черепно-мозговых нервов. Корковыми проекциями обонятельной
чувствительности являются структуры древней коры.
Двигательные области
коры расположены в прецентральной извилине лобной
доли (впереди от проекционных зон кожной чувствительности). Эту часть коры
занимают поля 4 и 5. Из V слоя этих полей берет начало пирамидный путь,
заканчивающийся на мотонейронах спинного мозга.
Расположение и соотношение зон иннервации сходно с проекционным представительством
кожного анализатора, т. е. имеет соматотопическую
организацию. В медиальных частях извилины расположены колонки, регулирующие
деятельность мышц ног, в нижней части, у латеральной борозды – мышц лица и
головы противоположной стороны тела.
Афферентные и
эфферентные проекционные зоны коры занимают относительно небольшую ее площадь.
Большая часть поверхности коры занята третичными или межанализаторными зонами, называемыми
ассоциативными. Ассоциативные зоны коры занимают значительное пространство
между лобной, затылочной и височной корой (60–70 % новой коры). Они получают
полимодальные входы от сенсорных областей коры и таламических ассоциативных
ядер и имеют выходы на двигательные зоны коры.
Ассоциативные зоны
обеспечивают интеграцию сенсорных входов и играют существенную роль в процессах
высшей нервной и психической деятельности.
Тема 8. Проводящие пути больших полушарий. Филогенез
и онтогенез конечного мозга. Черепные нервы. Возрастные
изменения коры больших полушарий
Белое вещество
конечного мозга составляет около 53 % полушарий, лежит между серым веществом
коры и подкорковыми ядрами и состоит из миелинизированных аксонов нервных клеток.
Волокна, идущие в одном направлении, объединяются в пучки и образуют структуры
белого вещества – проводящие пути.
Проводящие пути
больших полушарий подразделяют на проекционные, ассоциативные и комиссуральные.
Проекционные пути связывают
кору больших полушарий с нижележащими структурами. Те проекционные пути,
которые несут информацию от нижележащих структур в кору, называются афферентными
(в основном таламокортикальные). Проекционные пути, начинающиеся в коре и уходящие
к нижележащим структурам, являются эфферентными (например, пирамидные
тракты, идущие к спинному мозгу). Большая часть афферентных проекционных
волокон образована аксонами таламуса, восходящими к специфическим областям коры
– проекционным зонам. Выходя из таламуса, волокна объединяются в составе задней
ножки внутренней капсулы, отделяющей чечевицеобразное ядро от таламуса. Покидая
внутреннюю капсулу, афферентные волокна расходятся к проекционным зонам,
образуя лучистый венец, который состоит из отдельных радиаций: таламической,
слуховой, зрительной. Кроме того, по задней ножке внутренней капсулы проходят
проекционные эфферентные кортикоспинальные пути. По передней ножке внутренней
капсулы идут проекционные нисходящие (эфферентные) пути от лобной коры к
таламусу, красному ядру и ядрам моста. Через колено внутренней капсулы проходят
нисходящие пути к черепно-мозговым ядрам ствола. Таким образом, по внутренней капсуле проходят эфферентные корковые
проекционные пути и некоторые из афферентных путей.
Свод – эфферентный проекционный путь, связывающий кору гиппокампа с мамиллярными телами
гипоталамуса. Свод начинается от бахромок гиппокампа прилежащих к нему в области нижних рогов боковых
желудочков. Направляясь назад, они образуют ножки свода (рис. 44, 2).
Затем, поворачивая вверх и вперед, обе ножки сливаются в уплощенное тело свода
(рис. 44, 4). Тело свода идет продольно вперед над крышей
III желудочка под мозолистым телом. Боковые поверхности свода прилежат к таламусу.
Не доходя до колена мозолистого тела, свод круто изгибается вниз и вперед и
разделяется на два столба свода (рис. 44, 7). Каждый из столбов
идет латеральнее мозолистого тела, направляясь назад,
и образует часть латеральной стенки III желудочка. Заканчиваются столбы на
сосцевидных (мамиллярных) телах гипоталамуса.
Таким образом, свод
состоит из двух круто изогнутых дуг, каждая из которых начинается в нижней
части височной доли, направляется назад, вверх, медиально, соединяется со своей
парой в теле свода и, опускаясь вниз и латерально,
разделяется на столбы, оканчиваясь мамиллярными
телами. Кроме эфферентных проекционных путей, в состав свода входят
ассоциативные и комиссуральные волокна.
Ассоциативными путями называются
волокна, связывающие между собой различные участки коры одного полушария. Они
подразделяются на два типа – короткие и длинные.
Короткие, выходя из коры какой-либо извилины, дугообразно
изгибаются в подлежащем белом веществе вокруг борозды и заканчиваются в коре
прилежащих извилин.
Длинные ассоциативные волокна обеспечивают связь между долями
коры. Они, выходя из какой-либо доли, радиально собираются в пучки, идущие косо или изогнуто в белом веществе полушарий,
затем также веерообразно расходятся к коре другой доли.
Комиссуральные пути связывают между собой участки коры левого и правого
полушарий. Самой большой комиссурой является мозолистое тело. Оно
обеспечивает в основном связь между симметричными зонами новой коры и
представляет собой толстую белую спайку, большинство волокон которой идут в поперечном
направлении. В своем составе оно имеет до 200 млн волокон. Мозолистое тело
располагается в глубине продольной мозговой щели над сводом и крышей III мозгового
желудочка. Впереди тонкий нижний конец мозолистого тела граничит с концевой
пластинкой. Поворачивая вперед и вверх, он расширяется и выходит из-под
переднего отдела таламуса. Эта часть мозолистого тела называется клювом.
Клюв переходит в
сильноизогнутый участок – колено мозолистого тела. За коленом начинается ствол,
который, слегка изгибаясь над таламусом, идет назад. Затем следует изгиб вниз,
и мозолистое тело заканчивается расширенным закругленным участком-валиком, который
свободно нависает над передними отделами четверохолмия.
Поперечные волокна мозолистого тела, вступив в полушарие,
радиально расходятся, что можно наблюдать на горизонтальном срезе обоих
полушарий. Эти волокна образуют лучистость (радиацию) мозолистого тела. В
лучистости различают лобную, теменную и затылочную части соответственно долям
мозга, к которым направляются волокна. Волокна лучистости мозолистого тела
образуют крышу боковых желудочков.
Другой комиссурой является передняя спайка,
которая проходит под клювом мозолистого тела непосредственно у заднего края
концевой пластинки. В ее составе насчитывается до 3 млн волокон. По более
тонкой передней части спайки осуществляется связь структур древней коры. По задней
части (более широкой) спайки идут связи старой коры двух полушарий. Комиссура
свода, или спайка гиппокампа, связывает гиппокампы обоих полушарий. Волокна этой спайки
протягиваются между задними ножками свода.
Филогенез. У низших позвоночных (круглоротые и рыбы) развитие конечного мозга происходит
по пути утолщения основания, в котором формируются базальные ганглии в виде больших
парных выпуклых масс. Боковые стенки и крыша конечного мозга, образующие плащ
(мантию), остаются тонкими и повторяют форму полосатых тел в виде двух
полушарий, хотя на два полушария конечный мозг еще не разделен, не разделена
еще и полость его желудочка. В передней стенке конечного мозга развивается
парное выпячивание – обонятельные доли. Они продолжаются в более или менее
длинный стебелек, заканчивающийся обонятельной луковицей. Серое вещество мантии
выстилает полость желудочка, а ее поверхность содержит белое вещество. В
концевой пластинке располагается передняя комиссура, связывающая между собой
полосатые тела.
У двоякодышащих рыб и первых наземных позвоночных (амфибии)
передняя стенка мозгового пузыря вдвигается посередине между полосатыми телами
в виде продольной складки, разделяющей конечный мозг на два полушария. Полость
мозгового пузыря разделяется на два боковых желудочка, которые соединяются
сзади Монроевыми отверстиями с III желудочком. Мантия
еще тонкая, но нервные клетки перемещаются к поверхности, однако остаются под
слоем белого вещества. Над передней комиссурой появляется еще одна комиссура,
соединяющая между собой мантийные структуры полушарий (аналог комиссуры гиппокампа). У рептилий конечный мозг разрастается еще
больше, но это разрастание происходит в основном за счет развития базальных
ганглиев. Мантия остается тонкой, но на ее поверхности впервые появляется серое
вещество, которое представляет собой кору. Эта кора является высшей
обонятельной структурой и сравнима с гиппокампом
млекопитающих. Усложнение организации конечного мозга млекопитающих происходит
за счет развития мантии.
Черепные нервы (черепномозговые нервы) отходят от стволовой части головного мозга в
количестве 12 пар (обозначаются римскими цифрами по порядку их расположения).
I – обонятельные
нервы. Относятся к чувствительным нервам, образованы центральными
отростками биполярных обонятельных нейронов, которые располагаются в слизистой
оболочке обонятельной области полости носа. Обонятельные нервы не образуют
нервного ствола, собираются в 15–20 тонких нервов, которые проходят в
обонятельную луковицу. Волокна этих клеток входят в состав толстых обонятельных
путей, оканчивающихся латеральными и медиальными обонятельными полосками. По волокнам
этих полосок обонятельные импульсы попадают главным образом в древнюю кору, и
только часть волокон заканчивается в старой коре.
II – зрительные
нервы. Эта пара тоже относится к чувствительным нервам. Отдельный нерв
представляет собой толстый ствол, составленный из отростков ганглиозных нейроцитов ганглиозного слоя сетчатки глазного яблока. Зрительный
нерв начинается в сетчатке (в области слепого пятна), где отростки нейроцитов собираются в пучок (нерв). Нерв, прободая
сосудистую оболочку и склеру, проникает в полость черепа. Оба нерва (правый и
левый) образуют неполный перекрест (хиазму) и переходят в зрительные тракты.
III – глазодвигательные.
Данная пара относится к смешанным нервам. Одна часть начинается от
двигательного ядра, а другая – от вегетативного (парасимпатического) ядра. Ядра
располагаются в среднем мозге. Эти нервы осуществляют иннервацию почти всех
мышц глазного яблока.
IV – блоковые
нервы. Они относятся к двигательным нервам. Волокна их начинаются от ядра,
расположенного в среднем мозге. Нервы огибают ножку мозга далее через верхнеглазничную щель проникают в глазницу и иннервируют
верхнюю косую мышцу.
V – тройничные
нервы. Относятся к смешанным нервам. Их двигательные волокна начинаются от
ядра, расположенного в мосту. Чувствительные волокна подходят к ядрам
среднемозгового и спинномозгового пути тройничного нерва. Эта пара нервов иннервирует
кожу лица, лобную и височные области, слизистую оболочку носа, полость рта и
язык.
VI – отводящие
нервы. Это двигательные нервы. Они образованы аксонами двигательных
нейронов ядра отводящего нерва, расположенного в ромбовидной ямке. Они
иннервируют наружные прямые мышцы глаз.
VII – лицевые
нервы. Эта пара объединяет два нерва: собственно лицевой нерв
(двигательный) и промежуточный (чувствительный), следовательно, относится к
смешанной группе нервов. Ядра лицевых нервов залегают в пределах продолговатого
мозга и моста. Они иннервируют мимические мышцы лица и подкожные мышцы шеи.
Чувствительные волокна промежуточного нерва несут вкусовые импульсы от органа
вкуса.
VIII – преддверно-улитковые
нервы. Чувствительные нервы, образованы волокнами, идущими от нерва улитки
и нерва преддверия. Нерв улитки проводит импульсы от органа слуха, а нерв
преддверия – от вестибулярного аппарата.
IX – языкоглоточные
нервы. Смешанные нервы, образованные чувствительными, двигательными и
секреторными (парасимпатическими) волокнами. Чувствительные волокна иннервируют
вкусовые сосочки слизистой языка, глотки, а также слизистую среднего уха.
Двигательные волокна связаны с мышцами глотки.
X – блуждающие
нервы. Самые длинные из всех черепномозговых
нервов. Относятся к нервам смешанного типа. Отдельный нерв выходит из
продолговатого мозга. Блуждающие нервы имеют обширную область иннервации:
волокна, выходящие из вегетативного ядра, иннервируют органы грудной и брюшной
полости; другие волокна замедляют сердечный ритм и рефлекторно регулируют
уровень кровяного давления; часть волокон усиливают перистальтику кишечника, а
также оказывают суживающие влияние на бронхи, ряд волокон усиливают секрецию
желез желудочно-кишечного тракта.
XI – добавочные
нервы. Двигательные нервы. Они имеют два ядра, расположенные следующим
образом: одно находится в пределах продолговатого мозга, а другое – в спинном
мозге. Эта пара выходит в составе блуждающего и языкоглоточного нервов. Область
иннервации – грудино-ключично-сосцевидные и трапециевидные мышцы.
XII – подъязычные
нервы. Они являются двигательными нервами. Волокна начинаются от
двигательного ядра подъязычного нерва, расположенного в продолговатом мозге. Из
продолговатого мозга нерв выходит многочисленными корешками между пирамидой и
оливами. Данная пара нервов иннервирует все мышцы языка.
Онтогенез. В начальной стадии формирования мозга человека передний мозг является закругленным
концом нервной трубки. На этом этапе конечный мозг представлен тонкой ростральной
стенкой переднего мозга. Затем эта стенка выпячивается в дорсолатеральном направлении
и образует два мозговых пузыря. Полости этих пузырей образуют боковые желудочки.
На ростральном конце мозговой трубки и III желудочка находится участок ткани, соединяющий
оба пузыря конечного мозга. В дальнейшем он станет конечной пластинкой. Дно
мешка конечного мозга утолщается очень быстро, это утолщение образует полосатое
тело. В дальнейшем оно делится на хвостатое ядро, скорлупу, бледный шар и миндалину.
По мере роста полушарий базальные ядра смещаются медиально и примерно на 10-й неделе
развития (
Волокна, которые не заканчиваются в полосатом теле,
идут вдоль плоскости слияния основания конечного мозга с промежуточным, образуя
внутреннюю капсулу. Волокна, идущие каудально и огибающие снизу средний мозг,
образуют ножки мозга. Затем на вентральной поверхности продолговатого мозга они
образуют пирамиды. Уходя в спинной мозг, пирамидные пути перекрещиваются и
оканчиваются на мотонейронах спинного мозга.
Ассоциативные проводящие пути начинают выявляться в конце второго месяца
развития. Свод появляется из гиппокампа в конце
третьего месяца развития.
Мозолистое тело появляется в начале четвертого месяца
развития. Оно начинает формироваться в концевой пластинке в виде пучка
поперечных волокон, лежащих над комиссурой гиппокампа.
Мозолистое тело растет очень быстро в каудальном направлении и смещает туда же
комиссуру гиппокампа и свод. Гиппокамп
также смещается назад.
Пузыри конечного мозга разрастаются в ростральном, дорсальном
и каудальном направлениях. На ранних стадиях развития (до 3-го месяца) пузыри
имеют очень тонкую стенку и утолщаются медленно. Борозды и извилины начинают
формироваться только с 11–12-й недели. Первыми появляются латеральная и гиппокампова извилины. Затем процесс формирования борозд
протекает очень быстро. К моменту рождения существуют все основные извилины.
Возрастные изменения коры больших полушарий. В течение развития человека, начиная с
пятого месяца эмбриональной жизни до взрослого состояния, поверхность коры больших
полушарий увеличивается в 30 раз. Отдельные области коры до и после рождения
неодинаково, что связано с филогенетическим происхождением и их функциональными
особенностями. Помимо обонятельной сенсорной системы, связанной в основном с
древней корой, в новой коре раньше других приобретают признаки и свойства мозга взрослого человека корковые
концы двигательного и кожного анализаторов, а также лимбическая
система.
Лимбическая система связана с интерорецепторами,
регуляцией секрета слюнных эндокринных желез и поддержанием постоянства
внутренней среды организма. Затем дифференцируются корковые концы зрительного и
слухового анализаторов и ассоциативная верхнетеменная область, имеющая
отношение к тонкой кожной чувствительности – узнаванию предметов на ощупь.
При этом на протяжении
всего внеутробного развития относительная величина
затылочной области остается постоянной, составляя 12 % от всей поверхности
коры. Значительно позднее приближаются к строению мозга взрослого организма
лобная и нижняя теменная области, связанные с несколькими сенсорными системами.
У новорожденного лобная область составляет 20,6–21,5 % от всей поверхности
коры, тогда как у взрослого она равна 23,5 %. Нижнетеменная область у
новорожденного не превышает 6,5 %, у взрослого она равна 7,7 %.
Филогенетически наиболее
новые ассоциативные поля, называемые «специфически человеческими», имеющие
отношение к речедвигательному анализатору,
дифференцируются на более поздних этапах развития.
По мере развития ширина
коры большого мозга увеличивается в 2,5–3 раза по сравнению с периодом
новорожденности. Особенно интенсивно растут III слой коры и ассоциативные поля. В
течение всего периода развития наблюдается уменьшение числа нейронов на единицу
площади коры. Такое разреженное расположение нервных клеток объясняется
значительным ростом и увеличением размеров их отростков. Это приводит к раздвиганию
тел нейронов. Кроме того, различные области коры испытывают процесс миелинизации неодновременно. Первыми миелиновую обкладку
получают волокна проекционных путей. Позже явление миелинизации
охватывает наиболее молодые в филогенетическом отношении области коры большого
мозга.
Существенный скачок в своем
развитии и степени зрелости кора больших полушарий ребенка испытывает на 14-й
день после рождения. Особенно интенсивно увеличивается площадь полушарий
ребенка в первые два года жизни. Это объясняется процессом формирования сложных
целенаправленных движений и развитием речи. Дальнейшее качественное
совершенствование коры большого мозга наблюдается в 4 года и 7 лет, когда процессы
психической деятельности становятся разнообразней, сложней и богаче.
Строение и топография мозга
ребенка после 10 лет не имеет существенных различий по сравнению с мозгом
взрослого человека. Масса мозга ребенка в этот период составляет в среднем
Литература
1. Анатомия человека / под ред. М.Р. Сапина. – М.: Мир образования, 2002. – 430 с.
2. Андронеску А.
Анатомия ребенка. – Бухарест, 1970. – 397 с.
3. Bapyxa Э.А. Анатомия и эволюция нервной
системы. – Ростов н/Д, 1992. – 397 с.
4. Мозг, разум, поведение / под ред. Ф. Блума. – М.: Мир, 1989. – 432 с.
5. Сапин M.П., Билич Г.Л. Анатомия человека. – М., 1999. – 430 с.
6. Синельников Р.Д., Синельников
Я.Р. Атлас анатомии человека.
– М., 1989. Т. 3.
7. Шляхтин Г.В. Анатомия и эволюция нервной системы. – Саратов, 1984.
– 389 с.
Наида Муртазалиевна Абдуллаева
Магомед Магомедович Габибов
Людмила Мухаджировна Ортабаева
Анатомия центральной нервной системы
Курс лекций для студентов психологического факультета
Ч. II
Редактор Чипашвили Н.О.
Корректор Цахаева Э.И.
Компьютерная верстка Губарева
С.В.
Подписано в печать 16.03.2017.
Формат 60×84 1/16.
Усл. п. л. 4,38. Уч.-изд.
л. 3,23.
Тираж 100 экз. Заказ
Издательство ДГУ
г. Махачкала, ул. М. Ярагского, 59е