2.
Поперечно-полосатая мышечная ткань
Источником развития являются клетки миотомов миобласты. Различают головные, шейные, грудные,
поясничные, крестцовые миотомы. Они разрастаются в
дорзальном и вентральном направлениях. В них рано врастают ветви спинномозговых
нервов.
Часть миобластов
дифференцируется на месте (образуют аутохтонную
мускулатуру), а другие, с 3 недели внутриутробного развития мигрируют в
мезенхиму и, сливаясь, друг с другом образуют мышечные трубки (миотубы) с крупными центрально ориентированными ядрами. В миотубах происходит дифференцировка специальных органелл
миофибрилл. Первоначально они располагаются под плазмолеммой, а затем заполняют
большую часть миотубы. Ядра смещаются к периферии.
Клеточные центры и микротрубочки исчезают. Гранулярная ЭПС значительно
редуцируется. Такая многоядерная структура называется симпласт,
а для мышечной ткани - миосимпласт.
Часть миобластов
дифференцируется в миосателлитоциты, которые
располагаются на поверхности миосимпластов и
принимают участие в регенерации мышечной ткани.
Структурной единицей мышечной ткани является мышечное волокно, состоящее из миосимпласта и миосателлитоцитов, покрытых общей
базальной мембраной. Длина мышечных волокон колеблется от 1 до 40 мм, а толщина
0,1 мм.
В
мышечном волокне различают мембранный аппарат, фибриллярный (сократительный)
аппарат, трофический аппарат (ядро, саркоплазма, цитоплазматические органеллы).
Мембранный
аппарат. Каждое
мышечное волокно покрыто сарколеммой, где различают наружную базальную мембрану
и плазмолемму (под базальной мембраной), плазмолемма образует впячивания (Т-трубочки).
К плазмолемме
снаружи прилежат миосателлитоциты. При повреждении
базальной мембраны запускается митотический цикл миосателлитоцитов.
Фибриллярный
аппарат. Исчерченные волокна можно
разделить на составляющие их фибриллы (диаметром 1 мкм), названные
миофибриллами. В мышечном волокне они ориентированы продольно.
При
рассматривании мышечных волокон и миофибрилл под световым микроскопом,
отмечается чередование в них темных и светлых участков – дисков. Темные диски
отличаются двойным лучепреломлением и называются анизотропными дисками или А- дисками. Светлые диски не обладают двойным
лучепреломлением и называются изотропными или I – дисками. В средней части
диска А имеется более светлый участок Н-зона (участок
содержащий только толстые нити белка миозина). В области Н-зоны выделяется
более темная М-линия, состоящая из миомезина
(необходим для сборки толстых нитей и их фиксации при сокращении). В середине
диска I расположена плотная линия Z, которая построена из белковых фибриллярных
молекул. В частности, большую роль играет альфа актинин.
Z – линия соединена с соседними миофибриллами с помощью белка десмина и поэтому все названные линии и диски соседних
миофибрилл совпадают и создается картина
поперечно-полосатой исчерченности мышечного волокна.

Рис. 45. Саркомер.
Структурной единицей миофибриллы является саркомер (S) – это пучок миофиламентов
заключенный между двумя Z линиями (рис.45). Принимая во внимание вышеуказанные обозначения можно
структуру саркомера записать в виде формулы:
S= Z1 + 1/2 I1 +
А + 1/2 I2 + Z2
Под электронным
микроскопом миофибриллы представляют агрегаты из толстых (меозиновых)
филаментов (диаметр 14 нм,
длина 1500 нм, расстояние между ними 20-30 нм). Между толстыми филаментами
располагаются тонкие филаменты ( диаметр
7-8 нм).
Толстые филаменты
(миозиновые нити) состоят из молекул белка миозина. Он
является важнейшим сократительным белком мышцы. При непосредственном участии
миозина химическая энергия трансформируется в механическую работу. Каждая миозиновая нить состоит из 300-400 молекул миозина.
Молекула миозина – это гексамер, состоящий
из двух тяжелых и четырех легких цепей. Тяжелые цепи представляют собой две
спирально закрученные полипептидные нити. Они несут на своих концах глобулярные
(шаровидные) головки. Между головкой и тяжелой цепью – шарнирный участок, с
помощью которого головка может изменять свою конфигурацию. В области головок -
легкие цепи (по две на каждой). Молекулы миозина уложены в толстой нити таким
образом, что их головки обращены наружу, выступая над поверхностью толстой
нити, а тяжелые цепи образуют стержень толстой нити.
Тяжелые и легкие
цепи в молекуле миозина можно разделить обработкой мочевиной, гуанидинхлоридом и др. При мягкой обработке можно отделить
только легкие цепи. Миозину свойственна АТФ-азная
активность – высвобождающаяся энергия используется для мышечного сокращения.
Тонкие нити (актиновые
нити). Образованы тремя белками: актином, тропонином и тропомиозином.
Основным по массе белком является актин, который образует спираль. Молекулы тропомиозина располагаются в желобке этой спирали, молекулы
тропонина располагаются вдоль спирали.
Толстые нити
занимают центральную часть саркомера–А-диск, тонкие
занимают I диски и частично входят между толстыми миофиламентами. Только толстые нити содержит Н-зона.
При поступлении
нервных импульсов по аксонам двигательных нейронов происходит сокращение мышечного волокна.
Каждое мышечное волокно имеет собственный аппарат иннервации (моторная бляшка)
и окружено сетью гемокапилляров, располагающихся
в прилежащей рыхлой соединительной ткани. Этот комплекс называется мион. Группа мышечных волокон, которые иннервируются одним мотонейроном
называется нервно-мышечной единицей. Мышечные волокна в этом случае могут
располагаться не рядом (одно нервное окончание может контролировать от одного
до десятков мышечных волокон).
В покое взаимодействие тонких и толстых
нитей (миофиламентов) невозможно, т.к.
миозин-связывающие участки актина заблокированы тропомиозином.
При высокой концентрации ионов кальция конформационные
(пространственные) изменения тропомиозина приводят к
разблокированию миозин-связывающих участков молекул актина. Плазмолемма миосимпласта образует пальцевидные впячивания
(инвагинации) ориентированные поперечно по отношению к миосимпласту
называемые Т-трубочки. К каждой Т-трубочке примыкают по две цистерны саркоплазматического ретикулума
(гладкая ЭПС), образуя триаду: две цистерны и одна Т-трубочка. Са2+ концентрируется
в цистернах (там его концентрация в 800 раз больше, чем в саркоплазме).
Механизм
сокращения. При поступлении нервного импульса волна деполяризации доходит до цистерн
саркоплазматического ретикулума, из них выделяются
ионы кальция и концентрация кальция в саркоплазме
резко возрастает. Са2+ диффундирует к тонким нитям (филаментам) саркомера, где
связывается с тропонином и миозиновыми
головками. Это приводит:
1. К изменению конформации
(пространственного расположения) тропомиозина, что, в
свою очередь, приводит к освобождению участков актина, необходимых для
взаимодействия с миозиновыми головками.
2. Появлению АТФ-азной
активности миозина.
3. Взаимодействию миозиновых
головок с актином (актино- миозиновые «мостики») и
скольжению нитей (рис. 46).
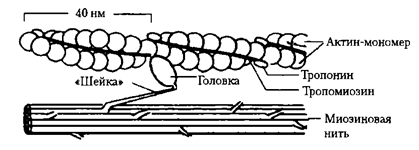
Рис.
46. Актомиозиновый
комплекс.
Все это вместе
взятое приводит к тому, что миозиновые головки
«шагают» по актину, образуя в ходе перемещения новые связи актина и миозина,
сближая две Z-линии. При сокращении уменьшаются только светлые диски.
Расслабление. Са2+-АТФ-аза
саркоплазматического ретикулума закачивает Са2+ из саркоплазмы в цистерны. В
саркоплазме концентрация Са2+ становится низкой. Са2+-тропомиозин
закрывает миозин-связывающие участки тонких нитей и препятствует их
взаимодействию с миозином.
Чувствительная иннервация (нервно-мышечные веретена). Интрафузальные мышечные волокна вместе с чувствительными
нервными окончаниями формируют нервно-мышечные веретена, являющиеся рецепторами
скелетной мышцы. Снаружи сформирована капсула веретена. При сокращении поперечно-полосатых (исчерченных) мышечных волокон
изменяется натяжение соединительно-тканной капсулы веретена и соответственно
изменяется тонус интрафузальных (расположенных под
капсулой) мышечных волокон. Формируется нервный импульс.
Классификация и типы мышечных волокон. Скелетные мышцы,
состоящие из мышечных волокон отличаются по многим параметрам: цвету, диаметру, утомляемости,
скорости сокращения и т.д. В каждой мышце присутствуют разные типы мышечных
волокон. В исчерченных мышцах различают два вида мышечных волокон: экстрафузальные, которые преобладают и обуславливают
собственно сократительную функцию мышцы и интрафузальные,
входящие в состав проприоцепторов–нервно-мышечных веретен.
По характеру
сокращения мышечные волокона делят на фазные и тонические.
Фазные способны осуществлять быстрые сокращения, но не могут длительно
удерживать достигнутый уровень укорочения. Тонические –обеспечивают
поддержание статического напряжения или тонуса.
По биохимическим
особенностям и цвету выделяют красные
и белые мышечные волокна. Цвет мышцы обусловлен степенью васкуляризации. Кроме того, существует прямая корреляция
между содержанием миоглобина и цветом мышцы. Характерной особенностью красных
мышечных волокон является наличие многочисленных митохондрий, цепи которых
располагаются между миофибриллами. В белых мышечных волокнах митохондрий меньше
и они располагаются равномерно в саркоплазме мышечного волокна.
Скорость
сокращения определяется типом миозина. Высокую скорость сокращения обеспечивает
быстрый миозин (для него характерна высокая активность АТФ-азы); меньшая
скорость сокращения характерна для медленного миозина (характерна невысокая
активность АТФ-азы). Следовательно, по активности АТФ-азы можно судить и о
наборе миозинов.
Тип
окислительного обмена.
Мышечные волокна используют два пути образования АТФ:
- при анаэробном типе метаболизма из 1
молекулы глюкозы образуется 2 молекулы АТФ и молочная кислота;
- при аэробном окислении из 1 молекулы
глюкозы образуется 38 молекул АТФ и конечные продукты метаболизма: СО2 и
Н2О.
Идентификация мышечных волокон основана на
выявлении активности фермента сукцинатдегидрогеназы
(СДГ), которая является маркером для митохондрий и цикла Кребса. Активность
этого фермента свидетельствует о напряженности энергетического метаболизма.
Выделяют мышечные волокна А-типа (гликолитические) с низкой активностью СДГ,
С-тип (оксидативные) с высокой активностью СДГ.
Мышечные волокна В-типа занимают промежуточное положение. Переход мышечных волоко от А-типа в С-тип
маркирует изменения от анаеробного гликолиза к
метаболизму, зависящему от кислорода.
Существует много
и других классификаций.
Факторами,
определяющими структуру и функцию скелетной мышцы являются влияние нервной ткани, гормональное
влияние, уровень васкуляризации, уровень двигательной
активности и местоположение мышцы.