Легочные объемы и емкости
Дыхательный
объем – это
объем воздуха, который вдыхает и выдыхает человек в спокойном состоянии; у
взрослого человека он равен 500 мл.
Резервный
объем вдоха – это
максимальный объем воздуха, который может вдохнуть человек после спокойного
вдоха; величина его равна 1,5-1,8 л.
Резервный
объем выдоха – это
максимальный объем воздуха, который может выдохнуть человек после спокойного
выдоха; этот объем составляет 1-1,5 л.
Остаточный
объем – это
объем воздуха, который остается в легких после максимального выдоха; величина
остаточного объема 1-1,5 л.
Жизненная
емкость легких (ЖЕЛ)
–
это максимальный объем воздуха, который может выдохнуть человек после самого
глубокого вдоха. ЖЕЛ включает в себя резервный объем вдоха, дыхательный объем и
резервный объем выдоха. Жизненная емкость легких определяется спирометром, а метод
ее определения называют спирометрией.
ЖЕЛ
зависит от:
1)
пола (у мужчин 4-5,5 л, а у женщин – 3-4,5 л)
2)
возраста
3)
натренированности (физическая тренировка приводит к увеличению ЖЕЛ.
4)
роста, веса
5)
от положения тела: она больше в сложении стоя, чем в положении сидя или лежа.
Функциональная
остаточная емкость (ФОЕ) – объем воздуха в легких после спокойного выдоха.
ФОЕ является суммой резервного объема выдоха и остаточного объема и равна 2,5
л.
Общая емкость
легких (ОЕЛ)
–
объем воздуха в легких по окончании полного вдоха. ОЕЛ включает в себя
остаточный объем и жизненную емкость легких.
Мертвое пространство
образует воздух, который находится в воздухоносных путях и не участвует в
газообмене. При вдохе последние порции атмосферного воздуха входят в мертвое
пространство и, не изменив своего состава, покидают его при выдохе. Объем
мертвого пространства 120-150 мл, или примерно 1/3 дыхательного
объема при спокойном дыхании. Значит, из 500 мл вдыхаемого воздуха в альвеолы
поступает лишь 350 мл. В альвеолах к концу спокойного выдоха находится около
2500 мл воздуха (ФОЕ), поэтому при каждом спокойном вдохе обновляется лишь 1/7
часть альвеолярного воздуха.
Тип дыхания зависит от рода трудовой деятельности и
пола. В детском возрасте отмечается грудной тип дыхания, после того, как
ребенок становится на ноги у девочек дыхание остается грудным, а у мальчиков
становится брюшным. При брюшном типе дыхания, в результате мощного сокращения
диафрагмы, органы брюшной полости смещаются вниз, поэтому при вдохе живот
«выпячивается». Грудной тип дыхания в значительной степени обеспечивается за
счет работы межреберных мышц. Однако доминирование того или другого типа
дыхания зависит главным образом от вида трудовой деятельности (вернее, от
положения тела при ее выполнении). Так, если выполнение физической работы
затрудняет грудное дыхание, то преимущественно брюшной тип дыхания формируется
и у женщин.
IV. Диффузия
газов в легких
В
обычных условиях человек дышит обычным воздухом, имеющим относительно
постоянный состав. В выдыхаемом воздухе всегда меньше кислорода и больше
углекислого газа. Меньше всего
кислорода и больше всего углекислого газа в альвеолярном воздухе. Различие в
составе альвеолярного и выдыхаемого воздуха объясняется тем, что последний
является смесью воздуха мертвого пространства и альвеолярного воздуха.
Альвеолярный
воздух является внутренней газовой средой организма. От его состава зависит
газовый состав артериальной крови. Состав альвеолярного воздуха при спокойном
дыхании мало зависит от фаз вдоха и выдоха.
Таблица 4.
Состав воздуха (в %)
|
Название газа |
Воздух |
||
|
вдыхаемый |
выдыхаемый |
альвеолярный |
|
|
Углекислый газ |
0,03 |
4,1 |
5,5-5,7 |
|
Кислород |
20,95 |
16,4 |
14,2-14,6 |
|
Азот |
79,02 |
79,5 |
80 |
Газообмен в
легких осуществляется в результате диффузии кислорода из альвеолярного воздуха
в кровь (около 500 л в сутки) и углекислого газа из крови в альвеолярный
воздух (около 430 л в сутки). Какие силы в легких заставляют кислород
устремляться в кровь, а углекислый газ – покидать ее?
Любой газ в
составе смеси (в данном случае такой
смесью будет вдыхаемый нами воздух) обладает собственной силой, называемой парциальным давлением (Р). Такой
же силой обладают и газы, растворенные в жидкой среде (кровь), только здесь эта
сила называется напряжением.
Обе силы измеряются в мм рт.ст. Вся «сцена» обмена разыгрывается в легочных
пузырях.
Если
парциальное давление газа в газовой смеси больше, чем напряжение этого же газа
в жидкости, газ стремиться проникнуть в жидкость и раствориться в ней, и
наоборот, если напряжение газа в жидкости больше его парциального давления в
газовой смеси, газ покидает жидкость.
На уровне
моря во вдыхаемом нами воздухе РО2 составляет около 100 мм рт.ст., а
его напряжение в венозной крови – 40 мм рт.ст. Естественно, кислород «давит» в
газе сильнее, чем «напрягает» в жидкости, и эта сила заставляет его поступать в
кровь, пока давление и напряжение кислорода не уравновесятся. Кровь протекает
через капилляры легких за 0,5 с, а чтобы кровь из венозной превратилась в
артериальную, достаточно половины этого времени. При здоровом состоянии
организма артериальная кровь насыщается кислородом на 95-97%.
Для
углекислого газа картина обратная. Его парциальное давление в альвеолах – 40 мм
рт.ст., а напряжение в крови – 46 мм рт.ст., поэтому СО2
выталкивается из крови пока не наступит равновесие. Несколько странным может
показаться тот факт, что, несмотря на меньшую разницу между напряжением и
давлением, СО2 покидает кровь
в 20 раз быстрее, чем О2 проникает в нее. Это происходит потому что
растворимость СО2 в 25 раз больше, чем О2. Тем не менее
артериальная кровь наряду с О2 всегда содержит небольшое количество
СО2.
Всю газовую смесь атмосферного воздуха принимают за
100%, она обладает давлением 760 мм рт. ст., а часть газа (кислорода – 20,95%)
принимают за х. Отсюда парциальное давление кислорода в смеси воздуха равно 159 мм рт.
ст. При расчете парциального давления газов в альвеолярном воздухе необходимо
учитывать, что он насыщен водяными парами, давление которых составляет 47 мм
рт. ст. Следовательно, на долю газовой смеси, входящей в состав альвеолярного
воздуха, приходится давление не 760 мм рт. ст., а 760 - 47 = 713 мм рт. ст. Это
давление принимается за 100%. Отсюда легко вычислить, что парциальное давление
кислорода, который содержится в альвеолярном воздухе в количестве 14,3%, будет
равно 102 мм рт. ст.; соответственно, расчет парциального давления углекислого
газа показывает, что оно равно 40 мм рт. ст.
Парциальное давление кислорода и углекислого газа в
альвеолярном воздухе является той силой, с которой молекулы этих газов стремятся
проникнуть через альвеолярную мембрану в кровь.
Если учесть, что в альвеолярной крови, притекающей к
легким, парциальное напряжение кислорода составляет 40 мм рт. ст., а углекислого
газа – 46-48 мм рт. ст., то градиент давления, определяющий диффузию газов в
легких, будет составлять: для кислорода 102 - 40 = 62 мм рт. ст.; для углекислого газа 40 -
46(48) = минус 6 - минус 8 мм рт. ст.
Времени протекания крови через капилляры
малого круга (в среднем 0,7 с) достаточно для практически полного выравнивания
парциального давления и напряжения газов: кислород растворяется в крови, а
углекислый газ переходит в альвеолярный воздух. Переход углекислого газа в
альвеолярный воздух при относительно небольшой разнице давлений объясняется
высокой диффузионной способностью легких для этого газа.
Таблица 5.
Парциальное давление и
напряжение О2 и СО2 в
легких (мм рт.ст.)
|
Название
газа |
Венозная кровь |
Альвеолярный воздух |
Артериальная кровь |
|
|
Кислород |
|
40 |
102 |
96 |
|
|
|
|
|
|
|
Углекислый газ |
|
46 |
40 |
39 |
V. Транспорт
газов кровью
Переносчиком кислорода от легких к тканям и
углекислого газа от тканей к легким является кровь. Кислород переносится с
кровью в растворенном виде (небольшое количество) и в соединении с гемоглобином
(основное количество кислорода).
Транспорт кислорода. Кислород, растворяющийся в плазме крови капилляров малого круга
кровообращения, диффундирует в эритроциты, сразу связывается с гемоглобином,
образуя оксигемоглобин. Скорость связывания кислорода велика: время
полунасыщения гемоглобина кислородом около 3 мс. Один грамм гемоглобина связывает
1,34 мл кислорода, в 100 мл крови 16 г гемоглобина и, следовательно, 19,0 мл
кислорода. Эта величина называется кислородной емкостью крови (КЕК).
Превращение гемоглобина в оксигемоглобин
определяется напряжением растворенного кислорода. Графически эта зависимость
выражается кривой диссоциации оксигемоглобина, или сатурационной кривой
(сатурация – насыщение жидкости газами) (рис. 28).
На рисунке видно, что даже при небольшом
парциальном давлении кислорода (40 мм рт. ст.) с ним связывается 75-80%
гемоглобина.
Кривая диссоциации имеет S-образную форму и состоит из двух частей –
крутой и отлогой. Отлогая часть кривой, соответствующая высоким (более 60 мм
рт. ст.) напряжениям кислорода, свидетельствует о том, что в этих условиях
содержание оксигемоглобина лишь слабо зависит от напряжения кислорода и его
парциального давления во вдыхаемом и альвеолярном воздухе. Верхняя отлогая
часть кривой диссоциации отражает способность гемоглобина связывать большие
количества кислорода, несмотря на умеренное снижение его парциального
давления.
При давлении 80-90 мм рт. ст. гемоглобин почти полностью насыщается
кислородом парциального давления во вдыхаемом воздухе. В этих условиях ткани
достаточно снабжаются кислородом (точка насыщения).
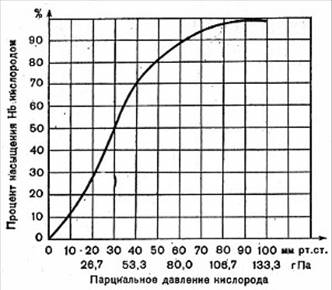
Рис. 29. Кривая диссоциации оксигемоглобина
Крутая часть кривой диссоциации соответствует
напряжению кислорода, обычному для тканей организма (35 мм рт. ст. и ниже). В
тканях, поглощающих много кислорода (работающие мышцы, печень, почки),
оксигемоглобин диссоциирует в большей степени, иногда почти полностью. В
тканях, в которых интенсивность окислительных процессов мала, большая часть
оксигемоглобина не диссоциирует.
Свойство гемоглобина – легко насыщаться
кислородом даже при небольших давлениях и легко его отдавать – очень важно.
Благодаря легкой отдаче гемоглобином кислорода при снижении его парциального
давления обеспечивается бесперебойное снабжение тканей кислородом, в которых
вследствие постоянного потребления кислорода его парциальное давление равно
нулю.
Распад оксигемоглобина на гемоглобин и
кислород зависит от:
1)
температуры (увеличивается с повышением температуры тела).
2)
реакции среды плазмы крови (с увеличением кислотности среды увеличивается
диссоциация оксигемоглобина)
3)
содержания солей (в воде связывание гемоглобина осуществляется быстро, но
полного насыщения не достигается, более полное насыщение кислородом и более
полная отдача кислорода происходит в физиологическом растворе).
4)
содержания в крови продуктов распада (способствуют более быстрой
диссоциации).
5)
содержания углекислого газа (чем больше содержание СО2 в
крови, тем меньше связывается гемоглобин с О2 и тем быстрее
происходит диссоциация оксигемоглобина).
Кривая диссоциации
оксигемоглобина смещается вправо при увеличении температуры, снижении рН и
повышении содержания 2,3 – ДФГ (капилляры большого круга кровообращения). Т.е в
этих условиях сродство гемоглобина к кислороду снижается. При увеличении рН,
снижении температуры тела и 2,3-ДФГ происходит сдвиг кривой влево (капилляры
малого круга кровообращения) (рис. 30).
Особенно
резко понижается способность гемоглобина соединяться с кислородом при давлении
углекислого газа, равном 46 мм рт. ст., т.е. при величине, соответствующей
напряжению углекислого газа в венозной крови. Влияние углекислого газа на
диссоциацию оксигемоглобина очень важно для переноса газов в легких и тканях.
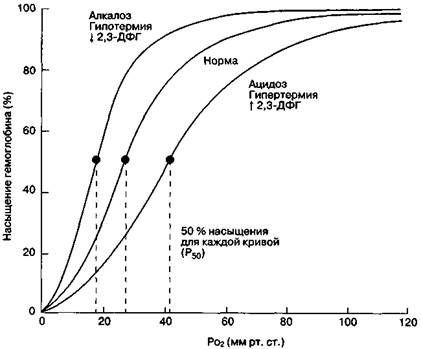
Рис. 30. Сдвиги кривой
диссоциации оксигемоглобина при изменениях рН, температуры тела и концентрации
2,3-дифосфоглицерата (2,3-ДФГ) в эритроцитах
 При подготовке к серьезным соревнованиям во время
разминки, даже в теплое время года спортсмены одевают утепленные костюмы
(разогревают мышцы). Зачем необходимо разогревать мышцы?
При подготовке к серьезным соревнованиям во время
разминки, даже в теплое время года спортсмены одевают утепленные костюмы
(разогревают мышцы). Зачем необходимо разогревать мышцы?
Транспорт
углекислого газа. Двуокись углерода переносится кровью
двумя основными способами:
1) физическим (растворением)
2) химическим.
Растворённый в плазме крови СО2
составляет около 5% общего количества транспортируемого кровью углекислого
газа. 95% СО2 переносится в форме химической связи с другими
веществами, содержащимися в крови.
Углекислый газ
находится в крови в одной из трёх химических форм:
1)угольная кислота (Н2СО3),
2) бикарбонатный ион
(НСО3-)
3) карбгемоглобин (HbCO2).
Связывание
СО2 кровью в капиллярах большого круга. Помимо
небольшого количества СО2, который попадая в кровь, физически
растворяется в её плазме, почти весь остальной углекислый газ, проникающий в
плазму капиллярной крови из тканевой жидкости сразу же диффундирует в
эритроциты, где осуществляются две основные химические реакции, связывающие СО2.
1-я
реакция - реакция
гидратации СО2 - связывание СО2 с водой с
образованием угольной кислоты (Н2СО3):
СО2 +
Н2О ⇄ Н2СО3
Эта реакция
катализируется ферментом – карбоангидразой;
участие фермента ускоряет реакцию в 250 раз. Как только образуется угольная
кислота, она немедленно ионизирует, т.е. диссоциирует на водородный ион (Н+)
и бикарбонатный ион (![]() ):
):
Н2СО3
⇄
Н+ + ![]()
Следовательно,
содержание угольной кислоты в крови ничтожно мало. Полностью 1-я реакция может
быть записана следующим образом:
СО2 + Н2О ⇄
Н2СО3 ⇄ Н+ +
![]()
Таким образом, в
результате 1-й реакции гидратации образуются бикарбонатные ионы, в составе
которых переносится более 4/5 всего содержащегося в венозной крови СО2.
По мере повышения концентрации бикарбонатных ионов в эритроците они
диффундируют в плазму, где соединяются с ионами Na+,
образуя бикарбонат натрия (NaНСО3).
Последний действует как буферный агент, предотвращающий повышение кислотности
(снижение рН) крови, особенно в связи с поступлением в кровь молочной кислоты (HLa) при мышечной работе.
HLa
+ NaНСО3
⇄NaLa + Н2СО3 ⇄
Н2О + СО2↑
2-я реакция:
связывание СО2 с белками крови и образование карбаминовых
соединений. Небольшая часть СО2 связывается прямо в плазме с её
белками. Однако в основном эта реакция осуществляется внутри эритроцитов, где
СО2 соединяется с гемоглобином, образуя карбгемоглобин. СО2
реагирует с глобином, не затрагивая частиц гема, которые сохраняют возможность
связи с О2.
Отдача
СО2 кровью в
лёгочных капиллярах. В крови альвеолярных капилляров наблюдается серия
противоположных по направлению реакций, которые происходят в основном в
эритроцитах и приводят к освобождению СО2 из химических связей и
выходу его из крови. Главная из них – реакция
дегидратации – совершается в эритроцитах с участием карбоангидразы:
Н+ + ![]() ⇄ Н2СО3
⇄
Н2О + СО2
⇄ Н2СО3
⇄
Н2О + СО2
Благодаря непрерывному
устранению конечного продукта (выходу СО2 из эритроцитов) реакция
идёт слева направо, как и другая реакция – диссоциация карбгемоглобина с
освобождением СО2 и присоединением О2 к восстановленному
гемоглобину:
О2 + H HbCO2⇄
H
HbO2
+ CO2
⇄
HbO2
+ Н+ + СО2
|
|
|

Рис. 31. Реакции, происходящие в процессе обмена кислородом и
углекислым газом между эритроцитом (I), плазмой (II) и
легочной альвеолой (III).