Лекции 4-5.
Тема:
Общая физиология ЦНС
План:
I.
Общая характеристика нервной системы
II.
Строение нейронов. Типы нейронов ЦНС.
III.
Физиология синапсов
IV.
Физиологические свойства медиаторов
V.
Рефлекторный принцип деятельности ЦНС и межнейрональные
взаимодействия. Классификация рефлексов.
VII.
Возбуждение и торможение в ЦНС.
I.
Общая
характеристика нервной системы.
С
помощью центральной нервной системы животный организм обеспечивает координацию
деятельности органов и систем, приспособление организма к изменениям окружающей
среды, формирует целенаправленное поведение и высшую нервную деятельность, связанную с психикой,
сознанием и памятью и т. д.
Еще
в 1891 году Вальдайер высказал мысль, что нервная система состоит из множества
нейронов, а не представляет собой синцитий – объединение клеток, не разделенных
мембраной. Далее работы Рамон - и - Кахала подтвердили нейронную теорию. Суть
теории состоит в том, что ЦНС - это функциональное объединение нейронов и
вспомогательных клеток глии. Позднее было показано, что обмен сигналами между
отдельными нейронами осуществляется с помощью синапсов.
Важнейшими
функциями нейронов являются восприятие, обработка хранение и передача
информации в форме электрических импульсов.
Мозг
человека насчитывает около 50 млрд нервных клеток. Нервные клетки имеются и вне
ЦНС – периферический отдел вегетативной нервной системы, афферентные нейроны
спинномозговых ганглиев и ганглиев черепных нервов.
Каждая
клетка ЦНС имеет множество синаптических контактов с различными нейронами. Так
число синапсов на одной клетке Пуркинье коры мозжечка доходит до 20 000
синапсов, на мотонейронах млекопитаюших –
от 10 000 до 20 000.
Схождение
различных путей проведения нервных
импульсов к одной и той же нервной клетке носит название конвергенция.
На клеточной мембране нейрона
конвергируют возбуждающие и тормозные входы, определяющие частоту
генерируемых им импульсов. В результате взаимодействия возбуждающих и тормозных
постсинаптических потенциалов обеспечивается интегративная деятельность нервной
клетки.
Нейроны
ЦНС способны устанавливать многочисленные синаптические связи с другими
нервными клетками. Это явление называется дивергенцией
и обнаруживается практически во
всех отделах центральной нервной системы.
Большая часть объема нервной
ткани (около 50%) составляют глиальные
клетки, или глия. Они сохраняют способность к делению в течение всей
жизни. Это деление определяет развитие опухолевых заболеваний головного мозга
(глиом). Число глиальных клеток достигает 14 10 в десятой степени.
В головном мозге выделяют три типа глиальных клеток:
1)
Микроглию – клетки участвуют в образовании мозговых
оболочек.
2)
олигодендроглию
- клетки участвуют в образовании миелиновых оболочек
3)
астроглию – клетки (астроциты) обеспечивают механическую,
трофическую и защитную функции нейронов.
II.
Строение нейронов. Типы нейронов ЦНС.
Нейроны (нейроциты, собственно нервные клетки) – клетки различных размеров
(которые варьируют от самых мелких в организме – у нейронов с диаметром тела
4-5мкм – до наиболее крупных с диаметром тела около 140 мкм). Их общее
количество в нервной системе человека превышает 100 млрд. (1011),
а по некоторым оценкам достигает одного триллиона (1012).
К рождению нейроны утрачивают способность к делению, поэтому в течение
постнатальной жизни их количество не увеличивается, а, напротив, в силу
естественной убыли клеток, постепенно снижается.
Гибель нейронов в физиологических условиях у взрослого
человека сравнительно невелика и
осуществляется механизмом апоптоза. Избыточной потере нейронов
препятствует их относительно высокая устойчивость к развитию апоптоза,
характерная для всех необновляемых клеток. Гибель нейронов значительно
ускоряется в старости, приводя к потере 20-40% клеток в некоторых участках
головного мозга.
Гибель нейронов при дегенеративных
заболеваниях нервной системы (болезнях Альцгеймера,
Гентингтона, Крейцфельда-Якоба,
паркинсонизме, боковом амиотрофическом склерозе и др.) осуществляется
вследствие ненормально высокой активности апоптоза, что приводит к резкому
снижению их содержания в определенных участках ЦНС. Развитие неврологических
нарушений, которые выявляются у 90% больных СПИДом, связано с потерей
40-50%нейронов в коре головного мозга, которые также погибают путем апоптоза.
В
каждой нервной клетке выделяют тело нейрона и его отростки. Тело нейрона
обеспечивает синтез макромолекул, которые транспортируются по отросткам, а
также трофическую функцию. Отростки нейрона представляют собой большое
количество дендритов и один аксон (рис. 19). Аксон начинается от тела клетки аксонным
холмиком (место генерации потенциала действия).
К специфическим
структурам нейрона относятся вещество Ниссля и нейрофиллменты.
Вещество Ниссля, или
тигроидное вещество представляет собой скопление уплощённых цистерн гранулярной
эндоплазматической сети, расположенных параллельно друг другу, которые при
окрашивании специальными красителями выявляются в виде базофильных зёрен или
глыбок. Характер
распределения и размеры комплексов цистерн грЭПС варьируют в отдельных типах
нейронов (наиболее крупные обнаруживаются в мотонейронах) и зависят от их
функционального состояния. При длительном раздражении или повреждении нейрона
комплексы цистерн грЭПС распадаются на отдельные элементы, что на
светооптическом уровне проявляется исчезновением телец Ниссля.
Нейрофиламентами
образуют трехмерную
опорно-сократительную сеть, играющую важную роль в поддержании формы этих
клеток и, в особенности, их длинного отростка – аксона. Нейрофибриллы составляют
опорную и дренажную систему нейронов. Не проводят нервных импульсов (нервные импульсы
проводит наружная мембрана).
Один
нейрон в результате многократного ветвления аксона, может образовывать синаптические
связи с 5 тыс. нейронов. На соме и дендритах заканчиваются синапсами аксоны от
различных нейронов. Возбуждение в нейроне проводится только от дендрита к
аксону. Диаметр тела нейрона составляет 10-100 мкм, аксона - 1 – 6 мкм, на
периферии длина аксона может достигать одного метра и более. У человека, аксоны
нейронов двигательной коры, так называемые клетки Беца, образующие
кортикоспинальный тракт, имеют около
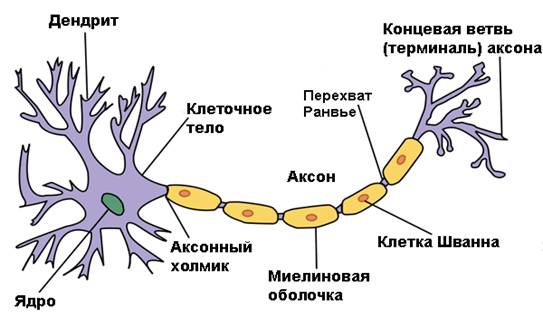
Рис.
19. Строение нервной клетки
Потенциал покоя нейрона составляет 60-80мв, потенциал действия –
80 – 100мв.
Классификация нейронов осуществляется по трем признакам: морфологическим, функциональным и биохимическим.
Морфологическая классификация нейронов учитывает
количество их
отростков и подразделяет все
нейроны на 4 типа (рис. 20): униполярные, биполярные, псевдоуниполярные и мультиполярные.
1.Униполярные нейроны имеют
один отросток. По мнению большинства исследователей, в нервной системе человека
и других млекопитающих они не
встречаются. Некоторые авторы к таким клеткам все же относят амакринные нейроны сетчатки
глаза и межклубочковые
нейроны обонятельной луковицы.
2. Биполярные
нейроны имеют два
отростка - аксон и
дендрит, обычно отходящие от противоположных полюсов клетки. В нервной
системе человека встречаются редко. К ним относят биполярные клетки сетчатки глаза,
спирального и вестибулярного ганглиев.
3.
Псевдоуниполярные нейроны – разновидность биполярных, в них оба
клеточных отростка (аксон и дендрит) отходят от тела клетки в виде единого
выроста, который далее Т-образно делится. Эти клетки встречаются в спинальных и краниальных
ганглиях.
4.Мультиполярные нейроны имеют
три или большее число от ростков: аксон и несколько дендритов. Они
наиболее распространены в нервной системе человека. Описано до 80 вариантов этих
клеток: веретенообразные, звездчатые, грушевидные, пирамидные, корзинчатые и
др. По длине аксона выделяют клетки Гольджи I типа (с длинным аксоном) и клетки Гольджи II типа (с
коротким аксоном).
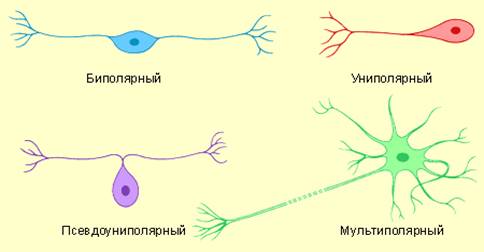
Рис.20. Морфологическая классификация нейронов.
Функциональная классификация нейронов разделяет их по
характеру выполняемой ими функции (в соответствии с их местом в рефлекторной дуге) на
три типа: чувствительные, двигательные и
ассоциативные.
1.Чувствительные (афферентные) нейроны генерируют нервные импульсы под влиянием
изменений внешней или внутренней среды.
2.Двигательные (эфферентные) нейроны передают сигналы на рабочие органы
(скелетные мышцы, железы, кровеносные сосуды).
3.Ассоциативные (вставочные) нейроны (интернейроны) осуществляют связи между нейронами и количественно преобладают над
нейронами других типов, составляя в нервной системе около 99.98% от общего
числа этих клеток.

Рис. 21. Функциональная
классификация нейронов
Биохимическая
классификация нейронов основана на химических
особенностях нейромедиаторов, используемых нейронами в синаптической передаче нервных импульсов.
1.
холинергические (медиатор - ацетилхолин),
2.
адренергические (медиатор -норадреналин);
3.
серотонинергические (медиатор - серотонин);
4.
дофаминергические (медиатор - дофамин);
1. ГАМК-ергические (медиатор - гамма-аминомаслянаякислота).
В некоторых нейронах терминали содержат
одновременно два типа нейромедиатора.
Синапс – место контакта двух возбудимых образований.
Термин "синапс" введен Ч. Шеррингтоном и означает
"сведение", "соединение", "застежка".
Синапс состоит из трех стуктурных образований:
1) пресинаптическая мембрана
2) постсинаптическая мембраны
3) синаптическая щель.
Конец аксона теряет миелиновую
оболочку и образует небольшое утолщение
(синаптическую бляшку).
Часть, контактирующая с иннервируемой клеткой, наз. пресинаптическая
мембрана.
Синаптическая щель – узкое
пространство между пресинаптической мембраной и мембраной иннервируемой клетки.
Постсинаптическая мембрана – участок
мембраны иннервируемой клетки, контактирующий с пресинаптической мембраной
через синаптическую щель.
В пресинаптическом окончании находятся синаптические
пузырьки (везикулы) диаметром до 200 нм. Они образуются в теле нейрона и с
помощью быстрого аксонного транспорта доставляются в пресинаптическое
окончание, где заполняются нейротрансмиттером, или медиатором (передатчиком). В
пресинаптическом окончании содержатся митохондрии, обеспечивающие энергией
процессы синаптической передачи. Эндоплазматическая сеть содержит депонированый
Са2+. Микротрубочки и микрофиламентыы участвуют в передвижении
везикул. Связывание Са2+с белками оболочки везикул приводит к
экзоцитозу медиатора в синаптическую щель.
Синаптическая щель имеет ширину от 20 до 50 нм,
содержит межклеточную жидкость и мукаполисахаридное плотное вещество для
обеспечения связей между пре- и постсинаптической мембранами, а также ферменты.
На
постсинаптической мембране синапса содержатся хеморецепторы, способные
связывать молекулы медиатора.
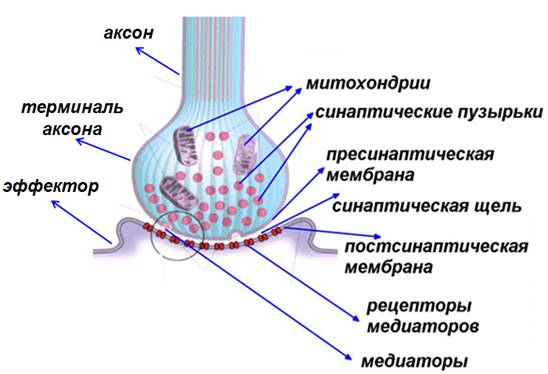
Рис. 22. Схема строения
синапса
Этапы передачи сигнала в химическом
синапсе следующие:
1) В пресинаптическое окончание поступает ПД.
2) ПД вызывает открытие потенциалчувствительных
кальциевых каналов в мембране пресинаптического окончания.
3) Через кальциевые каналы в пресинаптическое
окончание входят ионы Са2+.
4) Са2+ активирует белки, отвечающие за
слияние содержащих медиатор пузырьков с пресинаптической мембраной.
5) Слившись с мембраной, пузырьки путем экзоцитоза
высвобождают медиатор в синаптическую щель.
6) Молекулы медиатора взаимодействуют с рецепторами
на постсинаптической мембране и
активируют их.
7) Образование
постсинаптических потенциалов (потенциалов концевой пластинки – ПКП).
8) Суммация ПКП и возникновение на постсинаптической
мембране ПД.
Эти этапы характерны
для всех химических синапсов. Отличия же между разными синапсами касаются
следующих процессов:
- постсинаптической передачи сигнала, происходящей
после активации рецепторов.
- инактивации медиаторов.
- регуляции синаптической передачи.
Постсинаптические рецепторы
делятся на две большие группы (рис. 23):
1) Ионотропные рецепторы (сопряженные с ионными
каналами);
2) Метаботропные рецепторы (сопряженные с
внутриклеточными ферментативными системами).
Ионотропные рецепторы активируются
соответствующими медиаторами. Этапы передачи сигнала посредством таких
рецепторов следующие:
1)
Медиатор
активирует рецептор;
2)
Активация
рецепторов приводит к открыванию (реже – к закрыванию) ионного канала;
3)
Меняется
поток через канал ионов, для которых данный канал избирательно проницаем;
4)
Поток
ионов вызывает изменение заряда мембраны, возникает постсинаптический потенциал
- ПСП (потенциал концевой пластинки - ПКП). ПКП обладает всеми свойствами
местных потенциалов: распространяется с затуханием, подчиняется закону силы, не
обладает рефрактерностью, способен к суммации.
В зависимости от того,
для каких ионов проницаемы связанные с рецепторами каналы, могут возникать два
вида ПСП:
1)
Возбуждающие
(ВПСП)
2)
Тормозящие
(ТПСП).
Ионотропные рецепторы позволяют осуществлять гораздо
более быструю и точную регуляцию. Они используются при передаче возбуждения с
нейрона на нейрон или с нейрона на скелетную мышцу.
Метаботропные
рецепторы активируются не только медиаторами, но также гормонами и другими
биологически активными веществами. Они сопряжены с внутриклеточными
ферментативными системами: активация рецепторов непосредственно или через
промежуточные этапы приводит к активации фермента.
Метаботропные рецепторы обеспечивают более
длительный эффект, затрагивающий клетку в целом, ее функции и метаболизм. Они
используются в нервной регуляции внутренних органов.
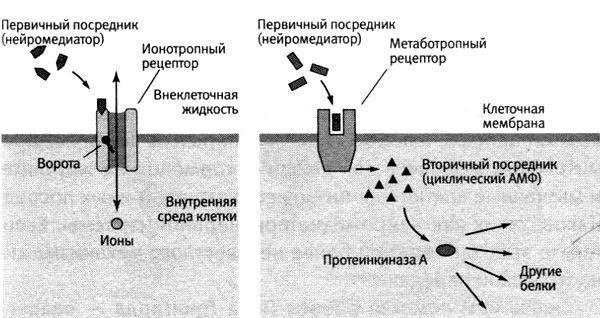
Рис. 23. Два типа рецепторов. Ионотропные рецепторы (слева)
вызывают изменения, которые длятся миллисекунды. Метаботропные рецепторы
(например, рецепторы серотонина) действуют через вторичных посредников
(справа). Они вызывают изменения, которые длятся секунды или минуты и
передаются по всей клетке.
После того как медиатор подействовал на рецепторы,
он должен быть удален из синаптической щели. Существует три способа этого
удаления, или инактивации медиатора:
1) Диффузия из
синаптической щели в окружающие ткани – самый простой, мдленный и неточный
способ (нервная регуляция сокращения кишечника)
2) Обратный захват пресинаптическим окончания –
более быстрый, экономичный способ.
3) Ферментативный распад медиатора – самый быстрый
способ, требующий присутствие фермента. Например, ацетилхолин разрушается под
действием ацетилхолинэстеразы на ацетил и холин.
Физиологические
свойства химических синапсов:
1)
Одностороннее проведение – от пресинаптической мембраны к постсинаптической;
2)
Синаптическая задержка. Это свойство обусловлено сравнительно длительным
временем, необходимым для выделения медиатора, его диффузии к рецепторам,
активации рецепторов и последующих постсинаптических процессов;
3)
Низкая лабильность. Синапсы становятся готовы к проведению очередного сигнала
лишь после того, как будет инактивирована очередная порция медиатора, на что
требуется много времени. Таким образом, лабильность синапса ограничена временем
полного оборота медиатора. Следовательно, синапсы способны проводить лишь
импульсы низкой частоты (максимум – 100-150 Гц), то есть обладают низкой
лабильностью.
4)
Высокая утомляемость. Причиной утомляемости синапса является исчерпание
ресурсов клетки, накопление метаболитов и пр.
5)
Происходит
трансформация ритма ПД.
Классификация синапсов ЦНС проводится
по нескольким признакам.
1)
По типу соединяемых клеток выделяют следующие синапсы:
- межнейронные, локализующиеся в ЦНС и
вегетативных ганглиях;
- нейроэффекторные (нейромышечные
и нейросекреторные), соединяющие эфферентные нейроны соматической и
вегетативной нервной системы с исполнительными клетками (поперечнополосатыми, и
гладкими мышечными волокнами, секреторными клетками);
- нейрорецепторные (контакты во вторичных
рецепторах между рецепторной клеткой и дендритом афферентного нейрона).
2) По типу контакта различают:
- аксосомаические,
- аксодендритные,
- аксоаксонные,
- дендросоматические,
- дендродендритные.
3) По функциональному эффекту
- возбуждающие;
- тормозящие.
4) По
типу медиатора:
- холинергические (медиатор – ацетилхолин);
- адренергические (норадреналин);
- дофаминергические (дофамин);
- ГАМК – ергические (гамма – аминомасляная кислота)
и т. д.
5) По способу передачи сигналов;
- химические синапсы, в которых
посредником (медиатором) передачи является химическое вещество;
- электрические, в которых сигналы передаются
электрическим током;
- электрохимические (смешанные).
Электрические
синапсы
были открыты в 1959 году Фершпаном и Поттером в брюшном нерве краба, а позднее
их обнаружили в многочисленных организмах. Электрический способ передачи сигналов в нервной системе
происходит между плотно прилегающими друг к другу клетками, образующими щель
размером всего около 2 нм. Например, между клетками миокарда и гладкой мускулатуры.
Столь плотный контакт обеспечивает возможность быстрой передачи электрического
импульса через ионные мостики-каналы. Подобными щелевыми контактами соединены
клетки эпителия и печени. Несмотря на быструю передачу возбуждения
(синаптическая задержка не превышает 0,1 мс по сравнению с 0,3 мс в химическом
синапсе), электрические синапсы имеют несколько недостатков:
– во-первых, с их помощью нельзя обеспечить
одностороннее проведение возбуждения,
– во-вторых, нельзя затормозить активность
эффекторной клетки,
– в-третьих, передача тока через электрический
синапс сопровождается потерей амплитуды.
- в-четвертых, преобразования (трансформации) ритма
возбуждения не происходит.
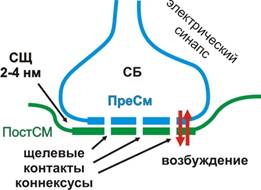
Рис.
24.
Строение электрического синапса
IV. Физиологические
свойства медиаторов.
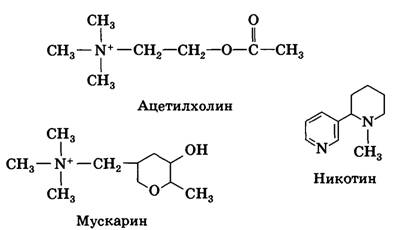
Ацетилхолин
является
достаточно широко распространенным возбуждающим медиатором в ЦНС. Он был открыт
в 30–х годах австрийским ученым О. Леви.
По химической природе ацетилхолин является уксуснокислым эфиром холина и
образуется при ацетилировании холина с участием фермента
ацетилхолинтрансферазы.
После
высвобождения из пресинаптических окончаний ацетилхолин быстро разрушается с
помощью фермента ацетилхолинэстеразы.
ацетилхолинтрансфераза
![]() Ацетил + холин
ацетилхолин
Ацетил + холин
ацетилхолин
![]()
Холин
является незаменимым витаминоподобным соединением, получаемым с пищей в
достаточном количестве, и последствия его дефицита наблюдаются только в
искусственных условиях. Синтез ацетилхолина осуществляется в основном в
пресинаптических окончаниях с помощью фермента холинацетилтрансферазы. Затем
медиатор переносится в пустые везикулы и хранится в них до момента выброса.
Ацетилхолин в качестве
медиатора работает в трех функциональных блоках нервной системы: в
нервно-мышечных синапсах, периферической части вегетативной нервной системы и
некоторых областях ЦНС (ретикулярная
формация ствола мозга, гипоталамус).
Выделяясь
из пресинаптического окончания, ацетилхолин действует на постсинаптические
рецепторы. Эти рецепторы неоднородны и различаются локализацией и рядом
свойств. Выделено два типа рецепторов:
1) Н-рецепторы
(никотиновые) - помимо ацетилхолина, возбуждается под действием алкалоида
табака никотина;
2) М-рецепторы
(мускариновые) - активируется ацетилхолином и токсином мухомора мускарином (мускариновые
рецепторы).
Никотиновые
рецепторы являются
классическим примером ионотропных рецепторов: их ионный канал входит в состав
рецептора и открывается сразу после присоединения ацетитилхолина.
Никотиновые рецепторы расположены на постсинаптической мембране
поперечнополосатых волокон скелетных мышц (нервно-мышечные синапсы); в синапсах
вегетативных ганглиев и в меньшем количестве, чем мускариновые рецепторы, в
ЦНС. Областью, наиболее чувствительной к никотину, являются вегетативные
ганглии, поэтому первые попытки курения приводят к значительным нарушениям в
деятельности
органов: скачкам артериального давления, тошноте, головокружению. По мере
привыкания сохраняется в основном симпатический компонент действия: никотин
начинает работать преимущественно как стимулятор многих систем организма.
Наиболее известным антагонистом никотиновых рецепторов является
тубокурарин - активное действующее начало яда некоторых южноамериканских
растений. Основным «местом приложения» его влияния являются нервно-мышечные
синапсы. При этом происходит последовательное расслабление и паралич мышц
пальцев, затем глаз, рук и ног, шеи, спины и, наконец, дыхательных.
В
настоящее время широко применяют синтетические аналоги кураре для
обездвиживания больных во время полостных операций в условиях искусственного
дыхания. Рецепторы к ацетилхолину в сердечной мышце – мускаринового типа и
кураре не останавливает сердце.
Эффект
действия ацетилхолина может быть как активирующим, так и инактивирующеим через
возбуждение тормозных интернейронов. Тормозное влияние ацетилхолин оказывает с
помощью М – холинорецепторов в глубоких слоях коры большого мозга, стволе
мозга, хвостатом ядре.
Мускариновые рецепторы являются метаботропными; они
связаны с G-белками, и присоединение к
ним ацетилхолина приводит к синтезу вторичных посредников.
Выделяют две основные локализации мускариновых рецепторов: синапсы,
образуемые постганглионарными (в основном парасимпатическими) вегетативными
волокнами и ЦНС.
Эффекты
мускарина носят преимущественно парасимпатический характер: при отравлении
мухоморами наблюдается тошнота, повышенное пото- и слюноотделение,
слезотечение, боли в животе, снижение артериального давления и сердечной
активности.
Классическим
антагонистом мускариновых рецепторов является атропин - токсин белены и
дурмана. Его периферические эффекты прямо противоположны действию мускарина:
происходит снижение тонуса мышц желудочно-кишечного тракта, учащается
сердцебиение, прекращается слюноотделение (сухость во рту), расширяются
зрачки, наблюдаются и центральные эффекты (двигательное и речевое возбуждение,
галлюцинации).
Нейроны
головного мозга, возбуждаемые через мускариновые ацетилхолиновые рецепторы,
играют большую роль в проявлении некоторых психических функций. Известно, что
гибель таких нейронов приводит к старческому слабоумию (болезнь Альцгеймера).
Биогенные
амины
включают две группы медиаторов: катехоламины (норадреналин, адреналин,
дофамин) и индоламин (серотонин).
Катехоламины являются производными
тирозина и выполняют медиаторную функцию в периферических и центральных
синапсах.
Норадреналин относится к катехоламинам – производным аминокислоты
тирозина. Тирозин - одна из незаменимых аминокислот, которые мы получаем
только с пищей. Цепочка химических превращений, ведущая к последовательному
образованию трех катехоламинов – дофамина, норадреналина и адреналина,
представлена ниже:
|
Тирозин |
|
L-Дофа |
|
Дофамин |
|
|
Норадреналин |
|
Адреналин |
|
|
|
Ключевая и наиболее медленная стадия - превращение тирозина в L-ДОФА
(диоксифенилаланин), где особое значение имеет регулирующий фермент тирозингидроксилаза.
Норадреналин играет важную роль в центральной и периферической нервной
системе.
На периферии норадреналин является медиатором большинства
постганглионарных симпатических синапсов. Воздействуя на внутренние органы, он
конкурирует с эффектами ацетилхолина. В ЦНС норадреналин вырабатывается нейронами
голубого пятна (мост) и межножкового ядра (средний мозг). Аксоны клеток этих
небольших ядер в дальнейшем можно встретить в различных структурах головного и
спинного мозга.
Синтез
норадреналина осуществляется в пресинаптических окончаниях, затем он
переносится в пустые везикулы и хранится до момента выброса. Выделяясь в
синаптическую щель, норадреналин действует на постсинаптические рецепторы, которые
неоднородны и подразделяются на два типа – альфа- и бета-адренорецепторы.
Оба они
являются метаботропными, но разница состоит в том, что в качестве вторичных
посредников эти рецепторы используют различные соединения.
Классическим
агонистом (вещество сходного действия) альфа-адренорецепторов является
препарат фетанол, антагонистом - фентоламин. В случае бета-адренорецепторов
наиболее известны агонист изадрин и
антагонист пропранолол.
В случае
симпатической нервной системы на каждом внутреннем органе можно встретить
альфа- или бета-адренорецепторы либо оба их типа. Альфа-рецепторы имеют
большинство сосудов, которые сужаются под действием норадреналина и агонистов
альфа-адренорецепторов, в результате чего повышается артериальное давление.
Агонистами альфа-адренорецепторов являются также препараты нафтизин (синоним -
санорин) и галазолин: при нанесении на слизистую носа они сужают сосуды,
прекращая насморк. Фентоламин, напротив, расширяет сосуды и снижает
артериальное давление.
Органами, содержащим только бета-адренорецепторы, являются сердечная
мышца и гладкие мышцы бронхов: норад-реналин и изадрин стимулируют деятельность
сердца и расширяют бронхи (последний эффект используется для ослабления
приступов бронхиальной астмы).
Функции норадреналина:
1) создание определенного уровня
активации бодрствующей ЦНС (за счет прежде всего торможения центров сна);
2) участие в тормозной регуляции
сенсорных потоков; обезболивающее (анальгетическое) действие, способное ярко
проявляться при сильном стрессе;
3) регуляция уровня двигательной
активности: норадреналин способен ее повышать, выключая тормозные интернейроны
в моторных центрах;
4) участие в регуляции
активности различных центров биологических потребностей и мотиваций (снижение
уровня тревожности, повышение уровня агрессивности);
5) влияние на выраженность
эмоциональных компонентов поведения: эмоции, возникающие в стрессовых условиях
(опасность, значительная умственная и физическая нагрузка), и эмоции,
соответствующие таким понятиям, как «азарт», «удовольствие от риска»; в
зависимости от индивидуальной организации мозга значимость таких эмоций для
конкретного человека может быть разной, но иногда - очень большой;
6) участие в процессах обучения
(запоминания информации), протекающих в корковых зонах ЦНС.
В отличие от ацетилхолина, катехоламины мало разлагаются в
синаптической щели, а в основном всасываются в пресинаптическое окончание.
Обратный захват норадреналина осуществляется особыми белками-насосами. Попав в
пресинаптическое окончание, норадреналин может повторно «загружаться» в
везикулы, но может и разлагаться с помощью фермента моноаминоксидазы (МАО). Инактивация происходит
внутри митохондрий, на внутренней мембране которых располагается МАО.
Чрезвычайно важно, что этот фермент осуществляет разложение и других моноаминов
— дофамина и серотонина. Оказалось, что использование блокаторов МАО позволяет
повысить активность всех трех медиаторных систем (антидепрессантные эффекты).
Дофамин встречается в
трех отделах головного мозга: черной субстанции, покрышке среднего мозга и в
различных ядрах гипоталамуса. В периферической нервной системе его практически
нет.
Нейроны
черной субстанции направляют аксоны к конечному мозгу, где образуют синапсы на
клетках полосатого тела. Функция этой проекции состоит в поддержании общего
уровня двигательной активности, обеспечении точности выполнения моторных
программ, устранении непроизвольных движений.
В
случае дегенерации черной субстанции наблюдается заболевание – паркинсонизм.
Его
основные симптомы состоят в:
1)
затрудненном
запуске движений (акинезия),
2)
патологически
усиленном мышечном тонусе (ригидность),
3)
дрожание пальцев и головы (тремор).
На
начальном этапе заболевания преобладает один из симптомов, позже они
комбинируются, образуя характерный комплекс двигательных нарушений.
Болезнь
обычно прогрессирует в течение 10-20 лет. Причины могут быть различны:
генетические аномалии, образовании токсических продуктов окисления дофамина.
Нарушение функции нейроглии и др. Риск развития паркинсонизма повышается с
возрастом (дагестанский писатель Расул Гамзатов), а также в результате
различных экстремальных воздействия на ЦНС: механические удары,отравления,
клиническая смерть (боксер Мухаммед Алли).
Препараты, применяемые для лечения
паркинсонизма, не устраняют его причин, а лишь облегчают тяжесть состояния,
ослабляют симптоматику. Они делятся на две группы. К первой относятся
антагонисты центральных никотиновых и мускариновых рецепторов (например,
циклодол). Снижая активность ацетилхолинергических интернейронов полосатого
тела, они способны значительно ослабить тремор.
Вторая группа препаратов — это L-ДОФА (L-диоксифени-лаланин) и его
производные.
L-ДОФА является
непосредственным предшественником дофамина в цепочке синтеза катехоламинов, а
также хорошо проходит гематоэнцефалический барьер. Превращаясь в дофамин
непосредственно в базальных ганглиях, он восполняет недостаток медиатора,
возникший в результате дегенерации черной субстанции. Влияние препарата
распространяется преимущественно на симптомы ригидности и акинезии. Однако,
действие его не продолжиельно.
Схема эффектов дофамина в норме и
патологии
|
|
В НОРМЕ |
|
|
Базальные
ганглии:
|
Кора больших полушарий:
|
Гипоталамус: гормональная регуляция |
|
|
ДОФАМИН
|
|
|
Недостаток в базальных ганглиях: паркинсонизм |
Избыток в коре больших полушарий: шизофрения |
Гормональные расстройства
под действием наркотиков |
|
|
ПРИ
ПАТОЛОГИИ |
|

Дофамин участвует в формировании чувства
удовольствия, регуляции эмоциональных реакций, поддержании бодрствования. Эффекты
дофамина в норме и при патологии представлены ниже.
Серотонин
(5 – гидрокситриптамин) наряду с катехоламинами относится к аминергическим медиаторам. Он образуется
путем гидроксилирования аминокислоты триптофана с последующим
декарбоксилированием. Химическая структура серотонина была расшифрована в 1952
году. 90% серотонина образуется в
организме клетками слизистой оболочки всего пищеварительного тракта. Большая
часть серотонина связывается с кровяными пластинками и по кровеносному руслу
разносится по организму.
Серотонинергические
нейроны широко распространены в центральной нервной системе, главным образом, в
структурах вегетативных функций. У человека он найден в различных отделах
головного мозга (мозговом стволе, варолиевом мосту, ядрах шва). С помощью
серотонина в нейронах ствола мозга передаются возбуждающие и тормозящие
влияния. Самая высокая концентрация серотонина обнаружена в эпифизе. Здесь
серотонин превращается в мелатонин, который участвует в пигментации кожи и
влияет на активность женских гонад.
Обязательным
условием для синтеза серотонина является наличие солнечного света, то есть,
если человек редко выходит на улицу или долго стоит пасмурная погода, то синтез
серотонина резко снижается, что приводит к снижению настроения. Поэтому
серотонин называют гормоном счастья, хорошего настроения.
Препараты,
повышающие настроение (антидепрессанты), блокируют захват серотонина в синапсах
после того, как он выполнил свою функцию.
Содержание
серотонина повышают также сладости, так как глюкоза вызывает выброс инсулина в
кровь, который стимулирует разложение белков на аминокислоты, в том числе и
триптофана.
Антагонистом серотонинорецепторов является
диэтиламид лизергиновой кислоты (LSD), который
является сильным галлюциногеном
|
|
свет |
|
темнота |
|
|
Триптофан (бананы,
черный шоколад, молочные продукты, рыба) |
|
серотонин |
|
мелатонин |
Физиологические эффекты серотонина связаны с
участием его в процессе обучения, формировании болевых ощущений, регуляции сна.
Серотонин играет важную роль в нисходящем контроле активности спинного мозга и
гипоталамическом контроле температуры тела. Нарушения функции
серотонинергических синапсов наблюдаются при шизофрении и других психических
расстройствах.
Аминокислотные медиаторы подразделяются
на две группы: возбуждающие (глутамат, аспартат) и тормозные (гамма –
аминомасляная кислота, глицин, бета – аланин и, таурин).
Глутамат в нервной ткани
образуется преимущественно из глюкозы.
Он является одним из самых распространенных медиаторов в ЦНС (75%
возбуждающих синапсов). Больше всего глутамата содержится в конечном мозге и
мозжечке, где его концентрация в 2 раза выше, чем в стволе мозга и спинном
мозге.
Будучи заменимой аминокислотой, он широко
распространен в самых разных белках. Однако, глутамат пищевого происхождения
очень плохо проникает через ГЭБ (гематоэнцефалический барьер), что предохраняет
нас от серьезных сбоев в работе мозга. Практически весь глутамат, необходимый
ЦНС, синтезируется в прямо в нервной ткани, но ситуация осложняется тем, что
данное вещество является также промежуточной стадией в процессах
внутриклеточного обмена аминокислот.
Однако, некоторая часть его может частично проходить
в тех зонах, где нет ГЭБ (гипоталамус и дно 4го желудочка). Взникающие при этом
изменения используют в клинике, назначая по 2-3 капли глутамата при задержке
психического развития, истощении нервной системы.
Кроме того, глутамат широко используется в пищевой
промышленности как вкусовая добавка (имеет мясной вкус). Очень богаты им
некоторые восточные преправы, изготовленные из морской капусты. Человек,
съевший несколько блюд японской кухни, может одномоментно получить 10-30 г
глутамата: последствием этого нередко становится активация сосудодвигательного
центра продолговатого мозга, рост артериального давления и учащение
сердцебиения.
Агонистами глутамата является каиновая кислота –
токсин одной из водорослей Японского моря. Вызывает судороги.
Антагонисты глутамата (ламотриджин, кетамин) в норме
оказывают тормозящее влияние на работу мозга и способны снижать патологическую
активность ЦНС. Препараты этой группы эффективны при эпилепсии, паркинсонизме,
болевом синдроме, некоторых видах депрессии.
Ламотриджин – перспективный противоэпилептический
препарат.
|
Глюкоза |
|
глутамат |
|
ГАМК |
Из тормозных медиаторов гамма – аминомасляная кислота (ГАМК) является самой
распространенной в ЦНС. ГАМК образуется из L
– глутаминовой кислоты путем декарбоксилирования. ГАМК обнаруживается в
мозжечке (клетки Пуркинье, клетки Гольджи, корзинчатые клетки), гиппокампе
(корзинчатые клетки), в обонятельной луковице и черной субстанции. Известно два
типа ГАМК рецепторов, которые связаны с хлорными каналами.
Агонистами ГАМК являются барбитураты и
транквилизаторы. Барбитураты – вещества, вызывающие генерализованное торможение
в ЦНС. Поэтому их используют как средства для наркоза, а также при тяжелых
формах эпилепсии как снотворное (барбитал). Транквилизаторы (лат. делать
спокойным) – вещества, снижающие эмоциональное напряжение, страх (элениум,
реланиум).
В последнее время особенно эффективным для
облегчения состояния больных эпилепсией считается совместное использование
агонистов ГАМК (бензодиазепинов) и антагонистов глутамата (ламотриджин). В этом
случае состояние нервной системы корректируется с двух сторон – за счет
усиления тормозящих и ослабления возбуждающих влияний.
Антагонистом ГАМК является бикукулин. Он хорошо
проходит через гематоэнцефалический барьер, оказывает сильное воздействие на
организм даже в малых концентрациях, вызывая
конвульсию и смерть. Содержится в семенах кукулевана.
Глицин. Вторым медиатором торможения,
которому приписывается существенная роль в работе мозга человека, является
глицин. В спинном и продолговатом мозге концентрация глицина достигает 3—5
мМ, но в коре
больших полушарий он содержится в небольшом количестве. Стрихнин служит специфическим антагонистом
рецепторов глицина в спинальных синапсах. Имеются данные, что действие столбнячного токсина обусловлено торможением высвобождения глицина из нейронов.
V. Рефлекторный принцип
деятельности ЦНС и межнейрональные взаимодействия. Классификация рефлексов
Рефлекс - ответная реакция организма на
раздражение сенсорных рецепторов, осуществляемая с помощью нервной системы.
Ответная реакция может быть в виде активации или торможения деятельности
органа. Каждый рефлекс осуществляется посредством рефлекторной дуги, состоящей
из нескольких звеньев.
Рефлекторная дуга - это совокупность структур, с помощью которых
осуществляется рефлекс. Схематично рефлекторную дугу вегетативного и
соматического рефлексов можно представить состоящей из пяти звеньев (рис. 21).
1) рецептор, обеспечивает восприятие
раздражителей (изменений внешней или внутренней среды организма), что
достигается посредством трансформации энергии раздражения в рецепторный
потенциал (РП). Совокупность рецепторов, раздражение которых вызывает рефлекс,
называют рефлексогенной зоной. Конкретные механизмы
восприятия раздражителя (света, звука, изменения химического состава крови)
различны у каждого вида рецепторов, но во всех случаях возникают РП, обеспечивающие
посылку нервных импульсов в ЦНС.
2) афферентный нейрон с отростками, его тело для
соматической нервной системы расположено в спинномозговых ганглиях или
ганглиях черепных нервов. Импульс поступает по дендриту афферентного нейрона,
а затем по его аксону - в ЦНС.
3) вставочные нейроны ЦНС.
Следует,
однако, отметить, что в составе ВНС вставочные (а также афферентные и
эфферентные) нейроны могут находиться вне ЦНС – интра- и/или экстраорганно. Их
назначение - обеспечение связи с другими отделами ЦНС, переработка и передача
импульсов к эффекторному нейрону.
4) эффекторный нейрон, он вместе с другими
нейронами ЦНС перерабатывает информацию, формирует ответ в виде нервных
импульсов, посылаемых к пятому
звену рефлекторной дуги - эффектору
(рабочему
органу).
5)
эффектор (рабочий орган) осуществляет действие.
В простейшем случае
рефлекторная дуга может быть двухнейронной – без вставочного нейрона (коленный
рефлекс).
Классификация рефлексов.
Выделяют соматические рефлексы, осуществляемые с помощью соматической
нервной системы (эффектором является скелетная мускулатура), и вегетативные рефлексы, осуществляемые с помощью ВНС.
По условиям появления рефлексов
в онтогенезе их подразделяют на две группы: врожденные (безусловные) и приобретенные (условные).
По биологическому значению
рефлексы делят на:
-гомеостатические
(регуляция
функций внутренних органов: работа сердца, секреция и моторика пищеварительной
системы - «пищевые» рефлексы и др.);
- защитные (оборонительные, например,
отдергивание конечности при ее раздражении);
- половые;
- ориентировочный рефлекс (рефлекс «что такое?») - непроизвольное
внимание к новому или внезапно появившемуся раздражителю (поворот головы, глаз,
туловища в сторону раздражителя).
В зависимости от числа синапсов в центральной части рефлекторной
дуги, различают моно- и полисинаптические рефлексы. Примером
моносинаптического рефлекса может служить рефлекс на растяжение четырехглавой
мышцы - коленный разгибательный рефлекс, возникающий при ударе по сухожилию
ниже надколенника. Большинство рефлексов являются полисинаптическими, в их
осуществлении участвует несколько последовательно включенных нейронов ЦНС, в
том числе расположенных на разных уровнях ЦНС.
По рецепторам, раздражение которых вызывает ответную реакцию,
различают экстероцептивные, интероцептивные и проприоцептивные рефлексы.
Последние используются в клинической практике для оценки состояния
возбудимости ЦНС и диагностики возможных ее поражений.
По локализации рефлекторной дуги различают:
- спинальные рефлексы (коленный,
локтевой, подошвенный);
- бульбарные (чихание,мигание, моргание, рвота, кашель);
- межэнцефальные (ориентировочный слуховой, зрачковый, сторожевой);
- корковые (условные рефлексы).
Нервный
центр - это совокупность нейронов, необходимых для осуществления определенного
рефлекса или регуляции той или иной
функции. Функционирование ЦНС в целом осуществляется благодаря
деятельности значительного числа таких центров, представляющих собой ансамбль
нервных клеток, объединенных с помощью синаптических контактов и отличающихся
многообразием и сложностью связей.
Свойства нервных центров зависят от
строения и механизмов передачи возбуждения в синапсах. К основным свойствам
нервных центров относятся:
1) Инерционность – сравнительно
медленное возникновение возбуждения всего комплекса нейронов центра при
поступлении к нему импульсов и медленное исчезновение после прекращения действия
раздражителя. Причины инерционности следующие:
1. Суммация возбуждения в ЦНС (пространственная и временная).
2.Последействие.
2) Трансформация ритма возбуждения.
Нервные центры способны изменять ритм,
приходящих к ним импульсов. Трансформация ритма возбуждения возможно как в
сторону увеличения, так и уменьшения. Уменьшение числа импульсов в нервном
центре происходит в результате снижения его возбудимости за счет пре– и
постсинаптического торможения, а также избыточным потоком афферентных
импульсов.
Трансформация частых ритмических
раздражений связана с низкой лабильностью синапса. При частом ритме
постсинаптические потенциалы суммируются, что приводит к стойкой деполяризации
постсинаптической мембраны и увеличению периода абсолютной рефрактерности.
Причиной трансформации ритма может быть
также вмешательство вставочных тормозных нейронов, блокирующих частый ритм еще
на пути к синапсу.
3) Фоновая активность нервных центров
(тонус) объясняется: 1) спонтанной активностью
нейронов ЦНС; 2) гуморальными воздействиями на биологическом уровне активных
веществ (метаболиты, гормоны, медиаторы и др.), циркулирующих в крови и
влияющих на возбудимость нейронов; 3) афферентной импульсацией от различных
рефлексогенных зон; 4) суммацией маленьких потенциалов, возникающих в итоге
спонтанного выделения квантов медиатора из аксонов, образующих синапсы на
нейронах; 5) циркуляцией возбуждения в ЦНС. Значение фоновой активности
нервных центров заключается в обеспечении некого начального уровня состояния
центра. Этот уровень может возрастать либо уменьшаться в зависимости от колебаний
суммарной активности нейронов нервного центра-регулятора.
4) Чувствительность к изменениям
внутренней среды.
Нейроны ЦНС чувствительны к изменению
содержания глюкозы в крови, газового состава крови, температуры, действию
фармакологических препаратов. При снижении содержания глюкозы до 50% от нормы
могут возникнуть судороги. Мозг чрезвычайно чувствителен к недлостатку
кислорода в крови. Прекращение кровотока на 8 – 12 мин вызывает необратимые
нарушения в деятельности мозга, гибели нейронов, в первую очередь корковых, что
ведет к тяжелым последствиям.
Нервные центры чувствительны к воздействию
некоторых фармакологических агентов.
4) Пластичность нервных центров.
Пластичность нервных центров проявляется в
способности нервных элементов к перестройке функциональных свойств. Основные
проявления этого свойства следующие.
1. Синаптическое облегчение – это
улучшение проведения в синапсах после короткого раздражения афферентгых путей.
Главной причиной возникновения синаптического облегчения является накопление
ионов кальция в пресинаптических окончаниях, увеличение высвобождения медиатора и возрастание ВПСП.
Синаптическое облегчение создает
предпосылки для улучшения процесса переработки информации на нейронах нервных центров.
Повторное возникновение явлений облегчения в нервном центре может вызвать
переход центра из обычного состояния в доминантное.
2. Образование временных связей,
обеспечивающих образование условных рефлексов. Этому способствует синаптическое
облегчение и доминантное состояние двух центров.
3. Доминанта – образование стойкого
господствующего очага возбуждения в ЦНС, подчиняющего себе функции других
нервных центров. На базе доминантного очага возбуждения формируется конкретная приспособительная
деятельность, ориентированная на достижение полезных результатов, для
устранения причин, поддерживающих тот или иной
нервный центр в доминантном состоянии.
4. Синаптическая депрессия
(утомление центра).
Утомляемость нервных центров связана с
расходованием медиатора при длительной активации синапса, а также накоплением
метаболитов и закислением среды.
5. Компенсация нарушенных функций.
После повреждения нервных центров возможна
компенсация нарушенных функций. Об этом свидетельствуют клинические наблюдения
за больными с нарушением тех или иных функций. Так при повреждении центров
регуляции мышечного тонуса и акта ходьбы, со временем отмечалось, что парализованная конечность у
больных.
VII. Возбуждение и торможение в ЦНС.
Возбуждение
и торможение в ЦНС обеспечивается функционированием возбуждающих и тормозных
химических синапсов. В возбуждающих синапсах осуществляется перенос нервного
импульса от одной клетки к другой. Примерами являются возбуждающее действие
центральных отростков первичных афферентых нейронов, мотонейронов и вставочных
клеток спинного мозга.
Для возбуждения нейрона необходимы поток
афферентных импульсов и их взаимодействие. Один, пришедший к нейрону
возбуждающий сигнал, сопровождается развитием локальной деполяризации на постсинаптической
мембране, обозначаемой как возбуждающий постсинаптический потенциал (ВПСП).
Возникновение ВПСП связано с одновременным увеличением проницаемости
постсинаптической мембраны для Nа+ и
К+, а также, возможно, Са2+. Амплитуда ВПСП составляет
всего 0,05 мв. Пороговый же потенциал, необходимый для генерации ПД составляет
– 5 – 10 мв. Поэтому для возбуждения нейрона требуется множество импульсов.
Наиболее эффективны синаптические контакты на теле нейрона, т.к.
постсинаптические мембраны этих синапсов располагаются в непосредственной
близости от аксонного холмика, места первичного возникновения ПД (генераторный
пункт нейрона). Отличительной особенностью мембраны аксонного холмика является
ее высокая возбудимость, отсутствие синапсов и миелиновой оболочки, высокая
концентрация Nа – каналов.
Впервые торможение в ЦНС открыл И.М.
Сеченов в 1863 году. В опыте на таламической лягушке он показал, что латентное
время сгибательного рефлекса при погружении задней конечности в слабый раствор
серной кислоты, значительно удлиняется, если на зрительный бугор (таламус) предварительно
положить кристаллик поваренной соли. Это открытие послужило толчком для
дальнейших исследований торможения в ЦНС.
Под
торможением в ЦНС понимают активный нервный процесс, результатом которого
является прекращение или ослабление возбуждения в ЦНС В настоящее время общепризнанно существование тормозных интернейронов.
Пресинаптические окончания аксонов тормозных нейронов образуют тормозные
синапсы. В тормозных синапсах полученный клеткой импульс сопровождается
подавлением другого возбуждения.
Примером тормозных нейронов в спинном мозге являются вставочные нейроны Реншоу,
в головном мозге - грушевидные нейроны Пуркинье коры мозжечка. Обнаружено два
совершенно различных механизма торможения: пресинаптический и
постсинаптический.
Постсинаптическое торможение. Этот вид
торможения имеет особенно широкое распространение в ЦНС. Вставочные нейроны
образуют тормозные синапсы на дендритах и телах мотонейронов, нервных клетках
мозгового ствола, гиппокампа и коры. Выделяемый здесь медиатор (глицин или
гамма – аминомасляная кислота) гиперполяризует постсинаптическую мембрану, т.е.
увеличивает мембранный потенциал этой области. Эта гиперполяризация получила
название тормозного постсинаптического потенциала (ТПСП).
Действие
тормозного медиатора при постсинаптическом торможении связано с увеличением
проницаемости постсинаптической мембраны мотонейрона для ионов калия и хлора.
Действие глицина на ионотропный рецептор постсинаптической мембраны сопровождается
увеличением ее проницаемости для ионов хлора. Ионы хлора поступают в клетку
согласно концентрационному градиенту, в результате чего развивается ТПСП. В
безхлорной среде тормозная функция глицина не реализуется.
При действии ГАМК на
постсинаптическую мембрану ТПСП развивается в результате входа С1- в
клетку или выхода К+ из клетки, что зависит от вида рецепторов, на
которые действует ГАМК. Имеются ГАМК-рецепторы двух типов: ГАМК1
(ГАМКА) и ГАМК2(ГАМКВ). Активация ГАМК1-рецепторов
(они ио-нотропные) ведёт к непосредственному повышению проницаемости клеточной
мембраны для CI-, который поступает в клетку,
как и при действии глицина, согласно концентрационному, но вопреки
электрическому, в результате чего развивается гиперполяризация и снижение возбудимости
нейрона. Активация ГАМК2-рецепторов (они метаботропные) реализуется
с помощью вторых посредников (цАМФ), при этом повышается проницаемость для К+,
который выходит из клетки тоже согласно концентрационному градиенту, но вопреки
электрическому, в результате чего также развивается гиперполяризация нейрона
(ТПСП), что снижает его возбудимость.
Существует несколько разновидностей
постсинаптического торможения: прямое, или реципрокное торможение и возвратное,
или антидромное торможение в спинном мозгу.
Понятие реципрокного торможения было
предложено Шерригтоном для объяснения работы мышц - антагонистов. Современное
представление об этом способе постсинаптического торможения было разработано
Экклсом (
Импульсы, идущие по афферентным волокнам,
отходящих от мышечных веретен (рецепторов растяжения), возбуждают рецепторы
своей мышцы (т.е. мышцы, от которой отходят афферентные волокна) и одновременно
через тормозные вставочные нейроны тормозят мотонейроны мышцы – антагониста.
Такие же отношения создаются между одноименными мышцами симметричных
конечностей.
Возвратное постсинаптическое торможение
осуществляется клетками Реншоу (тормозный медиатор – глицин). Аксоны
мотонейронов, идущие к мышцам, дают возвратную веточку в спинном мозгу,
которая оканчивается на клетках Реншоу. Аксон клетки Реншоу образует
тормозные синапсы на теле мотонейрона. Таким образом, мотонейрон при своем
возбуждении стимулирует клетку Реншоу, которая тормозит этот же мотонейрон. Это
торможение в центрах мышц-сгибателей и разгибателей обеспечивает поочередное
сокращение и расслабление скелетной мышцы, что необходимо при ходьбе и беге.
Пресинаптическое торможение. В основе механизма пресинаптического торможения лежит
деполяризация пресинаптических окончаний (тормозная пресинаптическая
деполяризация - ТПД). В очаге деполяризации нарушается процесс распространения
возбуждения, и, следовательно, поступающие к нервному окончанию импульсы, не
имея возможности пройти зону деполяризации в обычном количестве и обычной
амплитуды, не обеспечивают выделения медиатора из пресинаптического окончания
в синаптическую щель в достаточном количестве - пост-синаптический нейрон не
возбуждается, его функциональное состояние, естественно, остается неизменным.
Деполяризацию пресинаптической терминали вызывают специальные тормозные вставочные клетки, аксоны которых
образуют синапсы на пресинаптических окончаниях аксона-мишени. Снижение ПД в
пресинаптической терминали (частичная деполяризация) всего лишь на 5 %
уменьшает ВПСП на постсинаптической мембране на 50 %.
Торможение (деполяризация) терминали после одного
афферентного залпа продолжается 300-400 мс; медиатором служит ГАМК, которая
действует на ГАМК1-рецепторы. ТПД является следствием повышения
проницаемости для CI- и выхода его из клетки. Этот
факт свидетельствует о том, что в составе мембран пресинаптических терминалей
имеется хлорный насос, обеспечивающий первичный транспорт С1-
внутрь клетки вопреки электрическому градиенту. Под действием ГАМК тормозных
нейронов и последующего повышения проницаемости мембраны для С1-
ионы последнего начинают выходить наружу согласно электрическому градиенту,
но вопреки концентрационному. Это приводит к деполяризации пресинаптических
терминалей и ухудшению их способности проводить импульсы к постсинаптическому
нейрону. Роль ГАМК2-рецепторов на пресинаптических окончаниях
изучена недостаточно. Хотя известно, что активация ГАМК2-рецепторов
на постсинаптической мембране нейрона ведет к выходу ионов К+ из
клетки (гиперполяризация) и снижению возбудимости.
Роль
процессов торможения в ЦНС.
1) оба
вида торможения являются важным фактором обеспечения координационной
деятельности ЦНС. Об этом свидетельствует, в частности, тот факт, что блокада
торможения ведет к широкой иррадиации возбуждения и судорогам (например, при
выключении постсинаптического торможения столбнячным токсином).
2) оба
известных вида торможения, включая все их разновидности, выполняют
охранительную роль. Отсутствие торможения привело бы к истощению медиаторов в
аксонах нейронов и прекращению деятельности ЦНС.
3)
торможение играет важную роль в обработке поступающей в ЦНС информации.
Особенно ярко выражена эта роль у пресинаптического торможения. Оно более
точно регулирует процесс возбуждения, поскольку этим торможением могут быть
заблокированы отдельные синаптические входы. К одному возбуждающему нейрону могут
подходить сотни и тысячи импульсов по разным терминалям. Вместе с тем число
дошедших до нейрона импульсов определяется пресинаптическим торможением.
Оба механизма торможения широко
представлены в различных отделах ЦНС. ГАМК1-рецепторы
локализованы на нейронах гиппокампа, мозжечка, гипоталамуса, коры большого
мозга, на аксонах первичных афферентных клеток. ГАМК2-рецепторы
расположены в основном на терминалях моноаминер-гических нервных волокон и при
возбуждении тормозят секрецию медиатора. ГАМК повсеместно «сопровождает»
глутамат, прекращая его возбуждающее действие. Глицин - медиатор
постсинаптического торможения, кроме клеток Реншоу, обнаружен в стволе мозга.
Оба вида торможения могут быть заблокированы: пресинаптическое - бикукуллином,
постсинаптическое -стрихнином. Общим блокатором для постсинаптического и
пре-синаптического торможения является столбнячный токсин, который нарушает
высвобождение тормозных медиаторов.