К основным морфологическим и
функциональным особенностям приматов, выработанным в процессе эволюции в
условиях древесного образа жизни, относятся:
- хорошо развитая пятипалая
хватательная конечность, приспособленная к лазанию по ветвям деревьев;
- наличие развитой ключицы;
- полное разделение лучевой и
локтевой костей, многообразие движений: сгибание, разгибание, вращение лучевой
кости вокруг локтевой, позволяют производить супинацию и пронацию;
- подвижный большой палец,
который у многих видов может противопоставляться остальным;
- наличие когтей на концевых
фалангах пальцев;
- опора на стопу при
передвижении по земле;
- редукция обоняния и
усиленное развитие органов зрения и слуха;
- глазницы, окруженные костным
валиком и отделенные от височной ямы, обращены вперед, что обеспечивает
приматам бинокулярное зрение, имеющее важное значение в восприятии окружающей
среды в трехмерном пространстве;
- дифференцированный зубной
аппарат (4 типа зубов – резцы, клыки, малые и большие коренные зубы), имеет
место полная смена зубов;
- дерматоглифы по всей ладонной поверхности у обезьян и человека и только
на подушечках пальцев у полуобезьян;
- хорошо развитый головной
мозг, имеющий извилины у большинства приматов;
- наличие у всех приматов
плаценты, расположение семенников в мошонке;
- присутствие у самок двух
сосков и наличие овариально-менструального цикла;
- полицикличность размножения
без приуроченности к сезону;
- малая плодовитость (1–3
детеныша);
- забота о потомстве.
Все приматы – обитатели теплого климата:
Южная Америка, африканские леса и саванны, джунгли Индии, Малазии и леса
Мадагаскара.
Есть обезьяны, живущие на
снегу (Японский макак).
Ниже дана более подробная
морфологическая и функциональная характеристика некоторых семейств приматов.
3.1.2. Подотряд низшие приматы, или полуобезьяны
(Prosimii)
Подотряд низшие приматы, или полуобезьяны
(лемуры, тупайи, долгопяты и другие), включает 6 семейств (табл. 6).
В основном это мелкие животные,
но встречаются и более крупные, размером примерно с собаку (рис. 48).

Рис. 48. Полуобезьяны
Это хвостатые
обезьяны, хвост часто пушистый, лицевой череп вытянут, хорошо развито обоняние.
На лице располагаются осязательные волоски – вибриссы, нижние зубы растут
вперед и образуют гребешок для ухода за шерстью. Все они метят территорию
секретом желез или мочой. Мозг небольшой, без
извилин. Все полуобезьяны ведут ночной образ жизни, кроме некоторых видов
лемуров. Живут группами или поодиночке, мускулатура лица неподвижна за
исключением долгопятов, поэтому мимика у них не развита как у обезьян.
Семейство тупайеобразные (Tupaiidae). К этому семейству относятся
тупайи. Тупайи – переходная форма между
насекомоядными и приматами. По-малайски «тупайи» означает животных, похожих на
белку (рис. 48).
Характерные особенности
тупайи – длинное туловище и короткие
ноги. Большие пальцы не противопоставлены остальным, поэтому подвижность
конечностей ограничена. Глаза обращены в стороны, на мордочке присутствуют 4
пары вибрисс – чувствительных волосков. Мех
темно-бурый или рыжевато-коричневый. Мозг примитивный, лишен борозд и извилин.
Встречаются в горных лесах
(выше линии 3000 метров) Индии, Китая, Индокитая, на островах Суматра,
Калимантан и Бали.
Семейство лемуры (Lemuridae). Лемуры живут на Мадагаскаре. Свое
название получили от французского слова «Le mur» (мурлыканье), имеют густой
волосяной покров с разнообразной окраской. Хвост кошачьего лемура длинный и
волосатый (рис. 49) с чередующимися белыми и черными кольцами. Кошачий лемур
издает звуки, похожие на мурлыканье, за что и получил свое название. На
мордочке имеются 4–5 групп вибрисс, глаза большие, конечности хватательные,
большие пальцы противопоставлены остальным, все пальцы снабжены ногтями, а
второй палец имеет коготь, названный «туалетным», так как он служит для
расчесывания шерсти. Лемуры ведут дневной, ночной и сумеречный образ жизни. На
Мадагаскаре их около 30 видов. Есть и мышиные лемуры (50–60 г), они являются
ночными охотниками на насекомых.

Рис. 49. Лемуровые

Рис. 50. Долгопяты
Семейство долгопяты (Tarsiidae) включает 1 род Tarsius и 3 вида. Долгопяты получили свое название из-за
удлиненного пяточного отдела стопы (Tarsus). Эти животные небольшого
размера, стоят ближе к обезьянам, за огромные глаза, светящиеся в темноте, их
называют «долгопят – привидение». Передние
конечности короче задних, обе пары конечностей хватательного типа, на пальцах
имеются подушечки, используемые в качестве присосок при лазаньи по деревьям. На
всех пальцах имеются ногти, а второй и третий пальцы снабжены туалетными
коготками (рис. 50).
Долгопяты питаются
насекомыми, передвигаются прыжками в
Лицевой отдел укорочен, обоняние
слабое, хорошо развита мимическая мускулатура, гримасничают. Мозг сравнительно
большой.
Долгопяты распространены в
Индонезии и на Филиппинах.
3.1.3. Подотряд человекоподобные высшие приматы (Anthropoidea)
3.1.3.1. Надсемейство
широконосые обезьяны (Ceboidea)
Широконосые обезьяны обитают
только в Новом Свете (Америке). Для широконосых обезьян общими являются
следующие признаки. На пальцах обеих конечностей имеются плоские ногти. Глазницы
отделены от височной ямки сплошной костной перегородкой и находятся во
фронтальной плоскости; глаза направлены вперед. Мозг хорошо развит, большие
полушария у всех представителей покрыты бороздами и извилинами, за счет
редукции обонятельного мозга и строения носовых ходов обоняние слабое. Однако в
связи с дневным образом жизни у всех представителей человекоподобных, за
исключением некоторых обезьян Нового Света, хорошо развит орган зрения и на
сетчатке глаза имеются желтое пятно и центральная ямка.
В зубной системе общие черты
выражаются в наличии двух верхних и двух нижних резцов долотообразной формы.
Надсемейство широконосые
обезьяны объединяет два семейства: игрункообразные
(Callitrichidae) и цебиды (Cebidae).
Это в основном мелкие животные, отличаются примитивными чертами: мозг простой
почти без извилин (рис. 51), ушная раковина обволошена. Все игрунковые ведут
парный образ жизни. Размножается одна взрослая самка, а уход за потомством
осуществляет самец. Рождается до 3 детенышей.
Рис. 51. Мозг игрунка
Эти животные нашли применение
в экспериментальной медицине при изучении инфекционных заболеваний: гепатитов,
лейкозов и др.
В составе игрунковых выделено
в отдельное семейство коллимико – это мелкие обезьяны, которые по сходству
зубной системы, строению черепа и биохимическим показателям занимают
промежуточное положение между игрунками и капуцинами.
Семейство игрункообразные (Callitrichidae) –
обезьяны среднего размера, отличаются от игрунковых зубной системой (у них 36
зубов), длинным и хватательным хвостом, за исключением рода короткохвостых.
Мозг более развит, большие полушария несут борозды и извилины, поведение
сложное, их характеризует также яркая и разнообразная окраска.
Семейство цебиды (Cebidae) включает пять подсемейств:
коатовые, или паукообразные, капуцины, ночные обезьяны, ревуновые, чертовые
обезьяны, или саки.
Коаты интересны тем, что у
них длинный хватательный хвост с дерматоглифическим узором (рис. 52), а длинные
конечности придают им вид огромного паука. Они могут долго висеть, уцепившись
хвостом за ветку дерева (рис. 53). Передвигаются по земле, наступая полной
ступней и ладонью. Первый палец кисти редуцирован, а на ступне, наоборот,
противопоставлен.

Рис. 52. Кончик
хвоста коаты

Рис. 53. Коата (Atelesater)
Капуцинообразные, или
игрунки, собираются в сообщества для защиты от хищников, хорошо развиты
голосовое общение, мимика
Ревуны самые крупные из
широконосых обезьян. Они обладают плотным телом, на щеках растут густые волосы,
придавая им бородатый вид. Свое название они получили за громкий рев, которым
они наполняют воздух тропических лесов (рис. 54).

Рис. 54. Ревуны (Mycetesniger)
Чертовы обезьяны
характеризуются резцами, которые имеют сильный наклон вперед, а у одного вида –
саки – старый самец отличается
наличием бакенбардов и бороды.
3.1.3.2. Надсемейство собакоголовые,
или низшие узконосые обезьяны (Cercopithecoidea)
Надсемейство низшие узконосые
обезьяны (Cercopithecoidea) включает семейство мартышкообразные (Cercopithecoidae), в состав которого входят 8 родов.
Семейство мартышкообразные обезьяны. Представители данного
семейства распространены на обширной территории Старого Света: по всей Африке,
Индии, Аравии, в Индостане, Тибете, Японии, на островах Малайского архипелага,
Филиппинах, Тайване.
По размерам тела они крупнее
широконосых, весят в пределах от 3,5 до
Половой диморфизм хорошо
выражен, самцы крупнее самок. Передние конечности равны по длине задним, иногда
передние немного короче.
Первый палец обеих
конечностей противопоставлен остальным, хвост никогда не является хватательным,
окраска шерсти разнообразна в зависимости от вида, как правило, цвета яркие.
Лицо оголенное, ноздри в виде узких носовых щелей расположены не на конце
морды, а выше, и обращены вперед и вверх.
Характерной особенностью
мартышкообразных является наличие защечных мешков и седалищных мозолей.
Защечные мешки представляют собой карманы – складки слизистой оболочки ротовой полости,
куда набивается пища про запас. Седалищные мозоли прочно связаны с седалищными
буграми, которые обычно красного, желтого или синего цвета. Роль седалищных
бугров сводится к удобству во время сидения. Поза сидения хорошо усвоена всеми
приматами, однако в случае древесного образа жизни крупные обезьяны во время
сна могут сорваться с дерева, а мартышкообразные могут сидя спать на дереве. В
связи с этим седалищные мозоли обильно кровью не снабжаются, в них
откладываются подушечки жира в подкожной жировой клетчатке.
Кроме седалищных мозолей, для
мартышкообразных обезьян характерна «половая кожа», т. е. участок кожи, который
при очередной овуляции в результате прилива крови краснеет, что может быть
сигналом для самца о готовности самки к спариванию.
Все
представители низших обезьян ведут стадный образ жизни, к ним относятся роды мартышек,
гусар, павианов, мандрилов, гелад, мангобеев, макаков, колобусов, гверец,
лангуров и подсемейство тонкотелых обезьян. Они населяют леса, саванны и скалы.
Во многих арабских странах, в Индии некоторые виды обезьян причислены к
священным животным.
3.1.3.3.
Надсемейство человекоподобные приматы
(Hominoidea)
Надсемейство
человекоподобные приматы (Hominoidea) представлено
3 семействами, 6 родами, 12 видами.
Семейство
антропоморфных обезьян обладает характерными
признаками, сближающими их с человеком. Убедительные материалы, доказывающие их
генеалогическую близость с человеком, были представлены Томасом Гексли и
Чарльзом Дарвином. Сходства заключаются в морфологии, аппарате локомации и
различных системах органов. По внешним признакам сходство высших обезьян с
человеком выражается в крупных размерах тела, строении верхней губы, носа,
ушных раковин, коротком туловище при длинных ногах. Человекообразные обезьяны
передвигаются в выпрямленном состоянии по земле, а по деревьям – брахиацией, т. е. передвигаются с
помощью рук, перелетая с ветки на ветку или с дерева на дерево, тело при этом
находится в вертикальном положении.
Близость
антропоморфных обезьян к человеку выражается в больших размерах мозга (у гориллы
По
многим признакам внутренних органов также обнаруживается сходство антропоидов с
человеком. В частности, количество пирамид в почках у гориллы – 1–4, у шимпанзе
– от 1 до 6, а у человека варьирует от 3 до 20. Число долей в правом легком у
человека составляет 3, у гориллы и шимпанзе также 3, у низших узконосых их
число равно 4.
Существуют сходства также в строении гортани и языка.
Седалищные
бугры, характерные для низших обезьян, никогда не встречаются у человека, а из
высших антропоидов всегда присутствуют у гиббонов, чаще – у шимпанзе, а у
гориллы и орангутана встречаются значительно реже.
Поведение человекообразных
обезьян по сравнению с низшими гораздо сложнее. Условные рефлексы у шимпанзе
вырабатываются значительно быстрее, чем у макаки.
Особенно убедительны опыты с
распознаванием предметов. Шимпанзе легко распознает формы ящиков, где спрятана
пища, лучше и быстрее улавливает связь между тесемкой и приманкой, а низшим
обезьянам это не удавалось очень долго (Рогинский, 1978).
Неопровержимые факты
родственной близости человека и высших обезьян представлены сравнительной
эмбриологией. Противники эволюционной теории доказывали, что в отличие от всех
млекопитающих, у человека нет свободного аллантоидного пузыря, его заменяет
брюшной стебелек, заключающий в себе аллантоис в виде небольшого канала. У
человека оплодотворенное яйцо заключается в капсулу, образуемую слизистой
оболочкой матки. Однако позже исследователи (Зеленка, 1892–1902 гг., цит. по:
Рогинскому, Левину, 1978) доказали, что указанные особенности присущи и высшим
человекообразным обезьянам.
Для полноты общей
характеристики антропоидов следует остановиться на некоторых особенностях,
отличающих их как от низших обезьян, так и от человека.
У высших антропоидов
значительного развития достигают костные гребни на черепе самцов: у гориллы – надглазничный, сагиттальный и
затылочный; у шимпанзе – надглазничный.
Всеми высшими обезьянами
хорошо освоена брахиация, обеспечиваемая комплексом особенностей: передние
конечности длиннее задних, первый палец кисти и стопы несколько редуцирован
(при перепрыгивании с ветки на ветку он послужил бы помехой, а остальные четыре
пальца служат крюком при схватывании веток). Сухожилия глубокого сгибателя
кисти укорочены, что позволяет им висеть на дереве, не тратя силы на сгибание
кисти или стопы.
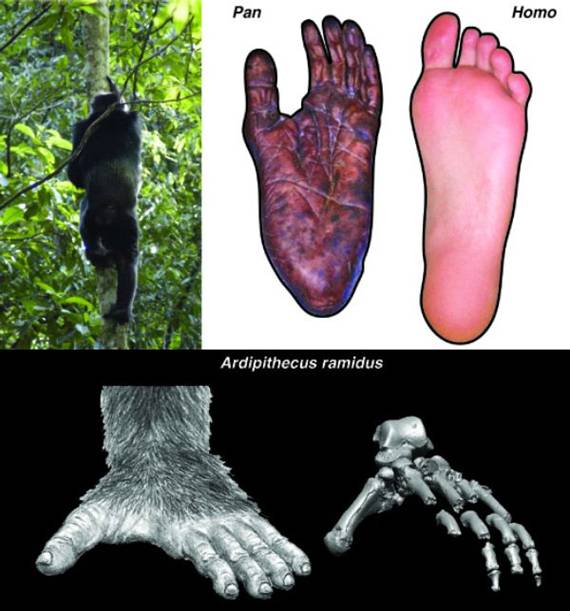
Рис. 55. Ступни современных
человекообразных обезьян специализированы для эффективного лазанья и хватания
за ветки, они очень гибкие и плохо приспособлены для ходьбы по земле. Слева
вверху: шимпанзе лезет на дерево; справа вверху: ступня шимпанзе и человека. У
ардипитека в строении ступни (внизу) наблюдается некоторые признаки,
свидетельствующие о сохранении способности хвататься за ветки и одновременно –
об эффективном двуногом хождении (более жесткий, чем у человекообразных
обезьян, свод стопы). Потомки ардипитеков – австралопитеки – утратили
способность хвататься ногами за ветки и приобрели почти человеческое строение
стопы. Изображение из статьи Lovejoy et. al. Combining Prehension and Propulsion: The Foot of
Ardipithecus ramidus (
Семейство гиббоновые (Hylobatidae)
включает 2 рода: гиббоны (Hilobata) и
сиаманг (Simphalangus) с 7 видами, распространенными в Юго-Восточной Азии и на
острове Суматра. Представители данного семейства относятся к малым
человекообразным обезьянам, занимающим промежуточное положение между высшими и
мартышкообразными обезьянами.
Гиббоны
отличаются наименьшими размерами тела среди высших обезьян, массой около
Головной мозг примитивен и
близок к мозгу низших человекообразных обезьян, его объем – около 120 см3.
Являясь обитателями
тропического леса, гиббоны прекрасно владеют брахиацией и способны перелетать с
дерева на дерево на расстояние до
Сиаманги, или сростнопалые
гиббоны (у них 2 и 3 пальцы стопы сращены), отличаются более крупными размерами
среди гиббонов и наличием горлового мешка, который служит резонатором для
усиления голоса.
Семейство понгиды (Pongidae) включает 3 рода (Pongo, Pan, Gorilla) с 4
видами. К антропоидам относятся шимпанзе и горилла, населяющие тропическую
Африку, орангутан и гиббоновые, живущие в Индии, Индокитае, на островах Ява,
Суматра, Калимантана и Малакка.
В лесах Калимантана и Суматры
встречаются крупные азиатские антропоиды – орангутаны (род Pongo). Орангутан
по-малайски означает «лесной человек». Для них характерен хорошо выраженный
половой диморфизм. Размеры самца достигают до

Рис. 56. Орангутан
Некоторые авторы выделяют два
подвида орангутанов: P. pigmaeus pigmaeus с острова Калимантана и P. pigmaeus abelii с острова Суматры.
Суматранские орангутаны по массе
тела (
Места обитания орангутанов –
болотистые леса, жизнь они проводят на деревьях, редко спускаясь на землю, так
как боятся и избегают воды. Передвигаются по деревьям брахиацией, но соблюдают
осторожность и не допускают перепрыгивания по веткам как гиббоны. Переходя с
одной руки на другую, предварительно проверяют устойчивость захвата. Несмотря
на медлительность, орангутаны достаточно быстро двигаются в вертикальном и
горизонтальном направлениях. При брахиации нижние конечности прижимаются к
туловищу, а тело сохраняет вертикальное положение. По земле двигаются на
четвереньках, опираясь на тыльную сторону пальцев и наружный край подошв стопы.
По наблюдениям Харрисона (
Самка вынашивает детеныша
около 40 недель, 3–4 года вскармливает молоком, одновременно воспитывает и
обучет передвигаться по деревьям и строить гнезда.
Род горилла (Gorilla) представлен одним видом
(Gorilla gorilla) с двумя подвидами: западная береговая горилла
(G. g. gorilla) встречается в Камеруне, Габоне, а восточная горная
горилла (G. g. beringei) имеет
небольшой ареал в горной области к северу и востоку от озера Киву (рис.
57).

Рис. 57. Горилла
Горилла является наиболее
крупным представителем приматов и превосходит размерами и массой тела всех современных
антропоидов. В среднем рост самца достигает 200 см, а масса тела – 200–250
кг. Половой диморфизм резко выражен. Горилла – это крупных размеров животное с
массивными туловищем и грудью, широкими плечами до 1 метра, большой
головой на короткой шее, мощной мускулатурой, обладает
огромной силой. Внешне имеет свирепый вид, хотя в целом является мирным
животным и на человека не нападает.
Глаза глубоко посажены под
мощным надбровьем, широкие ноздри окружены валиком, уши маленькие и прижатые к
голове, как у человека, лицо голое и черного цвета.
Верхние конечности гориллы
длинные, большой палец противопоставлен, несколько укорочен, но обладает
большой подвижностью. Кисть используется при строительстве гнезд. Первый палец
нижней конечности отставлен в сторону, а остальные пальцы соединены кожной
перепонкой вплоть до ногтевых фаланг. Тело покрыто короткой густой шерстью
черного цвета, только у взрослых самцов на спине имеется желтая или серебристая
полоса.
Горилла ведет стадный образ
жизни, в группе может быть от 5 до 30–40 особей. В группе существует иерархия,
состав сравнительно постоянный, однако она может пополняться за счет
присоединения новых самок с детенышами, а взрослые
самцы часто покидают стадо и примыкают к другим группам. В группе
взаимоотношения горилл в целом мирные, ссоры являются редким явлением, а вожак
с серебристой спиной чаще выступает как покровитель и не прибегает к жестким
наказаниям. Вожак не владеет большим гаремом, он не ревнив и половые отношения
не сопровождаются кровавыми поединками, они носят мягкий и добровольный
характер.
При соблюдении
последовательности движения по тропе или занятия более удобного места во время
отдыха или дождя на передний план выходят строгие иерархические отношения:
вожак идет впереди, остальные в соответствии с иерархическим рангом следуют за
вожаком.
Способ передвижения горилл по
деревьям и на земле, как у шимпанзе.
Гориллы строят гнезда для
ночного сна, иногда и для дневного отдыха. Повторное использование гнезд на вторую
ночь не происходит. После ночного сна гориллы пасутся и приступают к еде, они
питаются растительной пищей, в неволе могут питаться и животной пищей. Завтрак
длится 2 часа, после чего гориллы собираются вокруг вожака и отдыхают:
чистятся, чешутся, чаще это делают самки и подростки. Нередко во время дневного
отдыха вспыхивают ссоры между самками, которые сопровождаются воем, лаем,
иногда они визжат и кусаются, а вожак спокойно взирает на эти раздоры.
Гориллы сильные и могучие животные, они не ведают страха.
В редких случаях, когда ситуация становится опасной, вожак трясет ветку дерева,
бьет себя в грудь и издает крики.
Постоянных
мест для ночлега у горилл не бывает, поэтому, где их застает ночь, там они и
располагаются.
Размножение
горилл происходит круглый год, беременность длится 250–280 дней, рождается один
голый детеныш, который до 3 лет держится около матери, кормление грудью
заканчивается через год после рождения, а может длиться и дольше.
Продолжительность
жизни горилл в природных условиях составляет 30–35 лет.
Род
Шимпанзе (Pan) состоит из 2 видов: обыкновенный шимпанзе (Pan
Troglodites) и карликовый, или бонобо
(Pan Paniscus). Вид
обыкновенный шимпанзе подразделяют на три подвида, которые различаются по месту
обитания и пигментации лица. Подвид из Центральной Африки (Pttroglodites) отличается наличием на
белом фоне лица веснушек, которые с возрастом превращаются в более крупные
пятна грязноватого оттенка. Швейнфуртовский шимпанзе (P. t. Schweinfurthii), обитающий в Центральной и
Восточной Африке, отличается светлым лицом, переходящим с возрастом в темное и
грязноватое. Обыкновенный шимпанзе из западной Африки на лице имеет
бабочкообразную маску черного цвета, при этом надбровья и нижняя часть лица
остаются светлыми (рис. 58).

Рис. 58. Шимпанзе
Рост шимпанзе редко превышает
У шимпанзе верхние конечности
длиннее нижних, кисть длинная и узкая, 1 палец несколько укорочен. На стопах 1
палец крупный, а между остальными пальцами имеются кожные перепонки.
Нос у шимпанзе плоский,
верхняя губа оттопырена и участвует в мимике. Окраска шерсти черная, лицо, уши,
кисти и стопы голые.
Карликовый шимпанзе бонобо по
массе уступает обыкновенному почти в 2 раза.
Надглазничный валик развит
хорошо, а затылочный гребень отсутствует.
Шимпанзе обладает большой
силой. Опыты, проведенные с динамометром, показали, что он выжимает почти в
3 раза больше килограммов, чем средний атлет, очень проворен: с земли
может поднять иголку и взобраться на дерево с рюмкой, наполненной водой.
Шимпанзе ведет полуназемный и
полудревесный образ жизни, почти 30 % суток проводит на земле. Как и
орангутаны, они строят гнезда на дереве для ночного сна, а днем отдыхают на
земле или на дереве.
Питаются шимпанзе в основном
растительной пищей, однако в неволе могут употреблять и животную пищу:
муравьев, детенышей обезьян и других мелких животных.
Шимпанзе живут группами, в
составе которых может быть от 2 до 25 особей, иногда до 45 и больше.
Взаимоотношения в группах не строятся по правилам строгой иерархии.
Американский исследователь Д. Гудолл, наблюдая за ними в природных условиях,
отметила, что между взрослыми особями существует взаимное уважение, груминг.
Иногда между взрослыми подростками вспыхивают ссоры, которые быстро
прекращаются.
Размножение шимпанзе не
приурочено к определенному сезону. Беременность длится 225 дней, рождается один
детеныш, голый и беспомощный, он нуждается в уходе и длительном воспитании,
половая зрелость наступает у самок в 6–10 лет, а у самцов – в 7–8 лет.
Продолжительность жизни достигает 60 лет.
Семейство люди, или гоминиды (Hominidae), включает 2 подсемейства:
австралопитековые (Australopithecinae) и собственно люди (Homininae) с 1 родом
(Homo), 3 видами – человек умелый (H. habilis), человек прямоходящий
(H. erectus) и человек разумный (H. sapiens). Некоторые исследователи
считают, что состав семейства должен быть ограничен архантропами,
неандертальцами и современными людьми.
Исходя из изложенного
систематическое положение современного человека можно представить в виде следующей
схемы:
Отряд – приматы (Primates)
Подотряд – человекообразные (Anthropoidea)
Секция – узконосые (Catarrhini)
Надсемейство – высшие
узконосые (Hominoidea)
Семейство – гоминиды (Hominidae)
Подсемейство – люди (Hominidae)
Род – человек (Homo)
Вид – человек разумный (Homo sapiens).
Подсемейство современные люди (Hominidae) характеризуется ростом 140–190 см, самки на
10–12 см ниже. Передвижение осуществляется на нижних конечностях в выпрямленном
положении тела. Большой палец стопы не противопоставлен остальным и нижние
конечности длиннее верхних. Первый палец кисти противопоставлен остальным, что
позволяет человеку совершать разнообразные движения с помощью кисти.
Череп высокий и округлый, его
мозговая часть превалирует над лицевой. Лицевая часть не расположена впереди
мозговой, как у многих антропоидов, а находится под мозговой частью. Большое
затылочное отверстие находится снизу и посередине черепа. На жевательной
поверхности моляров верхней челюсти имеются четыре уплощенных бугорка, а на
нижней – по пять.
Позвоночный
столб имеет S-образную
форму с четырьмя изгибами: шейный и поясничный лордозы и грудной и крестцовый
кифозы. Позвоночник заканчивается копчиком из 4–5 сросшихся редуцированных
позвонков.
Головной
мозг человека хорошо развит, поверхность больших полушарий покрыта бороздами и
извилинами.
Беременность
продолжается 280 дней, рождается один ребенок, иногда два-три.
Характерной
особенностью человека, выработанной эволюцией, является наличие длительного
периода развития и воспитания ребенка.
Построение
филогенетической линии приматов опирается на материалы антропологических и
палеоантропологических исследований. В конце мелового периода мезозойской эры происходили
весьма значительные геологические и соответственно экологические процессы на
Земле, которые приводили к изменению биоты, населявшей наземную часть планеты.
В мезозойской эре, кроме того, происходило движение материков после «великого
раскола» Гондваны. Такой раскол сопровождался поднятием суши и обмелением морей
в северном полушарии и, наоборот, сокращением площади суши в южном полушарии.
Эти
изменения, вероятно, отразились на эволюционных процессах и привели к
возникновению примитивных приматов в конце мелового периода.
Для
ориентировки в геохронологии кайнозойской эры и эволюции приматов следует
обратиться к международной шкале подразделений, по Дж. Симпсону (
Таблица 7
Геохронологическая
шкала
|
Эра |
Период |
Продолжительность, млн
лет |
Эпоха |
|
Кайнозой |
Четвертичный (антропоген) Третичный (неоген) Палеоген |
1 24 35 |
Голоцен Плейстоцен Плиоцен Миоцен Олигоцен Эоцен Палеоцен |
|
Мезозой |
Меловой Юрский Триасовый |
70 45 40 |
Позднемеловой Раннемеловой Позднеюрский Среднеюрский Раннеюрский Позднетриасовый Среднетриасовый Раннетриасовый |
Предками приматов считают
древнейших насекомоядных плацентарных млекопитающих, сходных с современными
тупайями, населяющими Индо-Малайскую область.
Отделение приматов от
насекомоядных началось в конце мелового периода мезозойской эры 75 млн лет
назад.
В результате эволюционных
процессов у них прогрессивно развивался мозг, совершенствовались зрение,
осязание, повышалась функция хватания кисти и стопы, когти превратились в
ногти, усложнялся ладонный и подошвенный рельеф, связанный с тактильной
функцией. Однако зубная система у первичных приматов оставалась достаточно
примитивной, количество зубов составляло 44; зубная формула выглядела следующим
образом: ![]() , она
характерна для насекомоядных. Находка была сделана в Азии, в пустыне Гоби
(Монголия).
, она
характерна для насекомоядных. Находка была сделана в Азии, в пустыне Гоби
(Монголия).
3.2.1. Основные этапы
расцвета приматов в кайнозойской эре
По палеоантропологическим
данным в эволюции большинством исследователей выделяются три периода расцвета,
или радиации, связанные с возникновением приматов.
Первичная радиация приматов произошла
в раннем палеогене кайнозойской эры с формированием плезиантропов
(Plesiadapiformes) (рис. 59), которые по строению зубной системы и другим
признакам были сходны с современными полуобезьянами (лемурами и долгопятами).
Их было около 60 видов, все они были растительноядными и всеядными
животными.
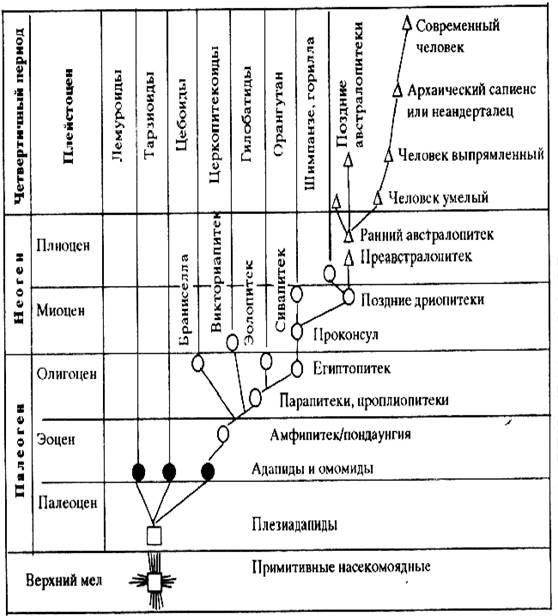
Рис. 59. Схема эволюции
приматов
В конце мезозоя в связи с
возникновением лесных массивов часть приматов приспособилась к лазанию по
деревьям, поэтому обе пары конечностей развились в хватательные с
противопоставлением первого пальца всем остальным. При лазаньи по деревьям
сохранение вертикального положения тела стало для них обычным и с течением
времени морфологические черты все больше приближали их к обезьянам.
Многообразные движения обезьян и высокая подвижность, свойственная древесным
млекопитающим, способствовали развитию двигательных участков головного мозга.
Растительная пища обогащала их организм необходимыми питательными веществами,
которые стимулировали дальнейшее увеличение размеров мозга. Переход этой группы
первичных приматов от насекомоядности к растительноядности и всеядности стал
вероятной причиной возникновения эволюционной линии, приведшей к полуобезьянам,
или лемурам.
Другая часть насекомоядных
приматов также приспособилась к древесному образу жизни: задние конечности в
связи с опорной функцией при передвижении прыжками или перепрыгивании с одного
дерева на другое стали длиннее и мощнее передних, а передними конечностями они
цеплялись за ветки деревьев и кустарников, т. е. осваивали передвижение
брахиацией – передвижение с помощью
рук. Кроме удлинения задних конечностей у них удлинялся пяточный отдел стопы,
что служило дополнительным рычагом для совершения очень больших прыжков с
дерева на дерево. По Земле они также передвигались прыжками.
Передними конечностями при
прыжках по деревьям необходимо было схватывать ветку, и в связи с этим на
передних конечностях оставались когти, кисть удлинилась за счет увеличения
размеров пальцевых фаланг.
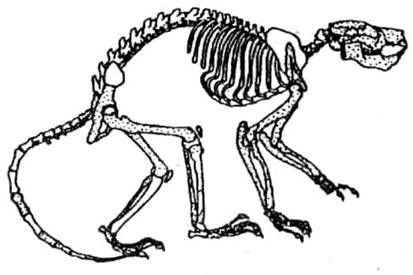
Рис. 60. Скелет плезиадаписа
Вероятно, от них берет начало
ветвь долгопятов, или тарзиоидов. Остатки скелета подобных животных встречаются
в раскопках Европы и Северной Америки (рис. 60).
Вторая
радиация первичных приматов произошла в самом начале эоцена кайнозойской эры,
она была связана с потеплением климата, обмелением северных морей, в результате
чего открылись мосты между Европой, Северной Америкой и Азией.
В Европе
и Северной Америке в это время появилась группа древнейших приматов,
относящихся к двум родам: тейярдина (Theilhardina) и кантиус (Cantius), которые представляли собой примитивные
древесные формы лемуроидов.
Полагают,
что они произошли от одного корня и являлись пришельцами из Азии или Африки.
Миграция их из Африки или Азии подвергается сомнениям, однако известно, что
полуобезьяны были широко распространены в Северной Америке, Европе, Азии и
Африке в течение эоцена. Их объединяют в две группы: адапиды (Adapidae), которых считают предками современных лемуроформов,
и омомиды (Omomyidae), которые являются предками тарзиоформов. Находки их
сделаны в Северной Америке и Европе.
На
рубеже эоцена и олигоцена произошел третий
расцвет приматов, который привел к появлению древнейших высших антропоидов,
или человекообразных обезьян. В связи с похолоданием и усилением конкуренции со
стороны грызунов или уже появившихся высших приматов большая часть ветвей
полуобезьян – омомидов и адапидов, относившихся к раннему
третичному периоду, вымерла. В этот же период возникают новые водные преграды,
прерывается связь между Северной Америкой и Европой, и при этом обезьяны
Старого и Нового Света развиваются своим эволюционным путем, независимо друг от
друга. Большинство исследователей считает, что омомиды, или ископаемые
долгопяты, являются предковой формой обезьян Старого и Нового Света, самые
ранние находки которых сделаны в Южной Америке (Бирме). Эти обезьяны получили
название амфипитеков и пондаунгия и сочетали черты полуобезьян
и обезьян, древность их определяется около 40 млн лет. Судя по строению зубов,
они были травоядными. Амфипитеки,
вероятно, являются предками узконосых обезьян. Тарзиоидная гипотеза выстраивается в линию: плезиадапиды →
омомиды → антропоиды.
Однако
существует гипотеза о происхождении высших приматов от лемуроидов, или адапид, – адапиоидная
гипотеза (Ф.
Гингрич).
3.2.2. Пути миграции приматов
в Южную Америку
Важное
значение в антропологии имеет вопрос о путях проникновения древнейших эоценовых
обезьян в Южную Америку. По этому поводу существуют разные гипотезы. Наиболее
вероятным считается «западный путь». По этой гипотезе проникновение обезьян
могло произойти через южную Атлантику, где существовала цепь островов,
соединявших Южную Америку с Африкой. Предполагают также существование
вулканических островов вдоль срединного Атлантического хребта, по которым могли
мигрировать приматы и грызуны в Южную Америку.
Гипотеза
«восточного пути» заселения Южной Америки азиатскими приматами предполагает,
что путь лежит через Берингию и Северную Америку, а дальнейшая их миграция
могла пройти через Панамский перешеек или Антильские острова и Никарагуанский
шельф.
Существует
также точка зрения автохтонного, или внутриконтинентального, развития
американских обезьян из Североамериканских полуобезьян. Однако ни одна из этих
гипотез не является строго доказанной.
В Южной
Америке сделаны самые ранние находки цебоидов:
брониселлы (Bronisella) из Боливии, долихоцебус (Dolichocebus) и тремоцебус (Tremocebus) из Аргентины. Все находки
относятся к олигоцену и представляют собой формы, сходные с современными
цебусовыми и игрунками.
Широконосые
обезьяны, самостоятельно развивавшиеся в Америке, адаптировались к жизни на
деревьях, развился цепкий хвост, в связи с разнообразным поведением увеличились
размеры головного мозга. Эти обезьяны связи с человеческой линией не имеют.
В Египте
сделаны находки узконосых обезьян олигоцена, они рассматриваются антропологами
как общие предки всех узконосых обезьян и человека – это парапитеки и проплиопитеки.
Эти
обезьяны сочетают в себе признаки строения полуобезьян (просимий), мартышковых
и человекообразных обезьян. Анализ строения тела и скелета показывает, что мозг
у них сравнительно большой, проплиопитеки адаптированы к лазанью по деревьям,
передвигались прыжками по деревьям и по земле на четырех ногах.
3.2.3.
Отделение группы ранних человекообразных
обезьян
От ранних узконосых обезьян отделились узконосые
человекообразные обезьяны, которые процветали в миоцене 23–20 млн лет назад в
результате экологической дивергенции. Подобные находки ранних антропоидов
обнаружены в миоценовых отложениях из Индии, Африки, Афганистана, Индонезии,
Китая. Плиопитек – одна из широко распространенных форм узконосых обезьян
считается предком гиббона. Они были хорошими брахиаторами, т. е. свободно
передвигались на руках, хватая ветки деревьев то одной, то другой рукой. В
отличие от современных гиббонов, характеризовались большей примитивностью по
строению челюстей и зубной системы.
Все
миоценовые антропоиды реконструируются в две филогенетические линии: азиатские
рамапитеки, давшие начало современным орангутанам, и дриопитеки, которые
являются предками шимпанзе и гориллы.
Рамапитек
– одна из наиболее важных
находок из миоценовых высших обезьян. Он был найден в Индии и назван в честь
персонажа из эпоса «Рамаяны» (рис. 61). Считалось, что рамапитеки достаточно
прогрессивные формы, они могли стоять на
двух ногах и использовать орудия.
Дриопитеки
интересны тем, что они представлены древнейшими останками антропоморфных обезьян,
кроме того, по некоторым признакам строения нижней челюсти и зубной системы они
больше сходны с человеком, чем любая из современных человекообразных обезьян.

Рис. 61. Dryopithecus (Proconsul) maior, самый крупный представитель
африканского рода Dryopithecus, достигавший размеров современной гориллы (Э.
Буриан)
Дриопитеки
были представлены двумя видами: дриопитек фонтанов и дриопитек дарвинов.
«Дриопитек» происходит от греческого слова «дриос» (дерево), они жили в Африке
и питались растительной пищей.
Ранние
дриопитеки найдены в Восточной Африке – Кении и Уганде. Среди них большой интерес
представляет африканский дриопитек проконсул,
находка которого сделана в 1933 г. у озера Виктория. В настоящее время эти
находки пополнены обширными палеонтологическими материалами, по которым
исторический возраст находок определяется в 24–16 млн лет.
В 60-е
годы ХХ в. в антропологии существовало мнение, что африканские дриопитеки могли
быть общим предком африканских понгидов и человека (так называемая «проконсулоидная гипотеза»).
Однако в
настоящее время такой взгляд пересмотрен, и дриопитеки рассматриваются как
примитивная группа ранних узконосых обезьян. По способу передвижения (древесная
и наземная локомация на четырех конечностях), пропорциям конечностей, строению
черепа они стоят ближе к низшим обезьянам, хотя степень церебрализации у них
выше. По экологическим приспособлениям они также более сходны с низшими
обезьянами, чем с шимпанзе.
Проконсулом
назвали ископаемую форму дриопитеков по кличке шимпанзе «Консул», жившего в
Лондонском зоопарке. Тем не менее группа ископаемых дриопитеков стоит ближе к
родословной человека, чем азиатские рамапитеки, которые представляют собой
боковую ветвь эволюции.
Способы
передвижения миоценовых обезьян многообразны: передвижение по деревьям и по
земле на четырех ногах, брахиация (передвижение по деревьям с помощью рук),
передвижение прыжками с опорой на задние ноги, лазанье по деревьям в вертикальном
положении тела. В результате движений на ногах усиливается таз и сочленения
костей задних конечностей с тазом, вероятно обезьяны также передвигались по
веткам на задних конечностях, придерживаясь руками за деревья. Такой способ
передвижения называется круриацией,
он считается многими исследователями исходной формой передвижения, которая
способствовала возникновению бипедии,
или хождения на двух ногах.
На
особом месте среди миоценовых антропоидов стоит семейство ореопитековых (Oreopithecoidea), живших в болотистых лесах Южной Европы. Они были
специализированными брахиаторами с элементами двуногого хождения и достаточно
крупным мозгом. Антропологи усматривают их генеалогическую связь с
олигоценовыми и раннемиоценовыми африканскими антропоидами.
Ореопитеки
считаются дриопитековой тупиковой ветвью эволюции приматов, однако их пример
указывает на возможность адаптации к бипедии уже в верхнем миоцене.
С 60-х
годов XX в. возможности исследователей по приматоантропогенезу значительно
расширились за счет внедрения в антропологию биомолекулярных методов,
т. е. развития нового направления восстановления филогенетических линий – молекулярных часов. Данные
молекулярной биологии совместно с палеонтологическими материалами позволяют
определить время дивергенции, или расхождения, гоминидной линии и
человекообразных обезьян. Подобная информация заложена в белках и
полинуклеотидах костной ткани вымерших приматов. За основу использования
молекулярных методов принят принцип, по которому мутации совершаются с неизменной
скоростью. Если учесть, что эволюция генетических структур (ДНК, РНК) и белков
происходит с постоянной скоростью, то структурные изменения гомологических
молекул разных видов животных соответствуют времени,
прошедшему с момента их эволюционной дивергенции. На рисунке 62 представлены
периоды дивергенции отдельных групп приматов по данным биомолекулярных часов.
Время
расхождения гоминидной линии и африканских понгид (шимпанзе и горилла)
соответствует 6 млн лет, а мартышкообразных – 25 млн лет.
Анализ
палеонтологических материалов не позволяет выявить бесспорное исходное звено
человеческой линии эволюции в миоценовую эпоху, тем не менее, согласно наиболее
распространенной точке зрения, эволюция человека заняла 10 млн лет и
вероятным предком его был антропоид типа шимпанзе (шимпанзиодная теория). По молекулярным данным человек входит в один
кластер с шимпанзе.
Следует
оговориться, что нельзя представлять предка человека копией одного из
современных видов человекообразных обезьян.
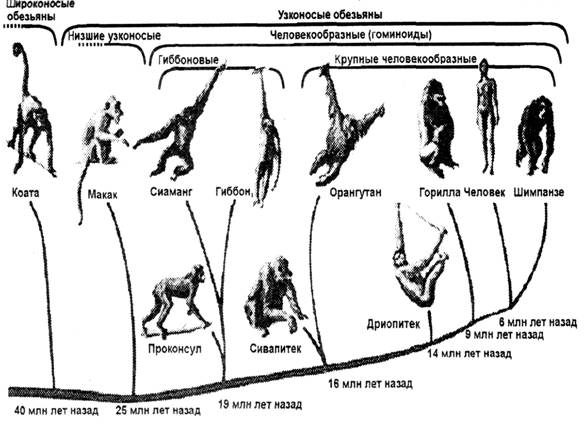
Рис. 62. Дивергенция
некоторых групп приматов по данным
биомолекулярного
анализа
Обзор находок гоминид миоценовой эпохи показывает, что выделить
среди них несомненного предка человеческой линии эволюции не удается. В связи с
этим «переходное звено» между человеком и обезьяной нужно было искать на
границе третичного и четвертичного периодов в плиоценовую и плиоплейстоценовую эпохи, когда жили двуногие
гоминиды – австралопитеки (Australopithecus – australis
«южная», pithecus «обезьяна» (рис. 63).

Рис. 63. Australopithecus
africanus
3.3.1. Австралопитек африканский
История
изучения австралопитеков началась в 1924 году с находки черепа гоминида.
Находка была сделана в Южной Африке Трансваале (ныне ЮАР) близ Таунга. Раскопки
велись под руководством профессора Раймонда Дарта (

Рис. 64. Череп Australopithecus africanus
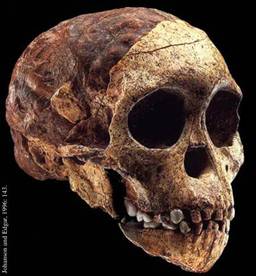
Рис. 65. Череп детеныша Australopithecus africanus
В том же
блоке был найден почти полный череп, принадлежавший детенышу, которому было не
более 6 лет, с полным набором молочных зубов. Р. Дарт назвал находку «Бэби из
Таунга» (рис. 65). Судя по тому, что большое затылочное отверстие располагалось
снизу черепа, это был прямоходящий гоминид. Объем мозга Бэби составлял 520 см3.
Надглазничные
валики, характерные для обезьян, отсутствовали, клыки не выходили из соседнего
зубного ряда, как у обезьян. Развитие отдельных зон мозга свидетельствовало о
сложном поведении «Бэби». Исторический возраст находки был определен Дартом в 1 млн лет, по сведениям других авторов, его
возраст около 3 млн лет. Научное название «Бэби» – австралопитек
африканский (Australopithecus africanus).
В
пещерах Южной Африки вместе с остатками австралопитека африканского обнаружено
много разбитых черепов павианов. Р. Дарт полагал, что они охотились на павианов
и убивали их камнями и дубинами. Позже К.К. Брейн установил,
что австралопитеки африканские были не охотниками, а, скорее всего, жертвами
(Lee R. Berger, 2006). Какие-то хищники (саблезуб, пятнистая гиена, леопард)
убивали павианов, а австралопитеки затаскивали их в пещеры. В настоящее время
многие специалисты считают, что австралопитеки не умели охотиться, а питались
падалью или остатками пиршества хищников. Изучение изотопного состава эмали
показало, что они употребляли и растительную и животную пищу.
В последующие годы Дарт совместно с палеонтологом
Р. Брумом сделали ряд других находок австралопитековых в Южной Африке: в
Стерхфонтейне, Макапансгате, Кромдраае. Их остатки находили в пещерах в толще
карбонатных и доломитных отложений. Находкам давались родовые названия:
плезиантропы (Plesianthropus),
парантропы (Paranthropus), однако, по современным
представлениям, среди южно-африканских австралопитековых выделяют один род с
двумя видами: древнюю, или изящную, форму называют грацильными (A. africanus), а более позднюю – массивными (A. robustus) (рис. 66–68).

Рис. 66. Paranthropus robustus

Рис. 67. Череп Paranthropus robustus

Рис. 68. Paranthropus robustus
В 1947–1949 гг. Дарт в породе пещеры
Макапансгат обнаружил следы пепла, фрагменты черепов и костей таза, зубы
австралопитеков, которых за следы огня он назвал австралопитеком прометеевым. Здесь были найдены «кухонные кучи»
австралопитека, где встречались кости разных животных: антилоп, павианов,
черепах, птиц и др. Исходя из своих находок, Дарт предположил, что
австралопитеки охотились на животных и даже применяли орудия для охоты и
разделки туш. В пещере было найдено более 50 черепов со следами ударов. Дарт
считал, что австралопитеки наносили удары по голове павианов тяжелыми
трубчатыми костями антилоп. Вмятины от ударов находились на левой стороне черепа,
что свидетельствует, по мнению Дарта, о праворукости австралопитеков. Дарт
назвал использование костей и рогов животных для охоты и раздела туш
«костно-зубо-роговой культурой».
3.3.2.
Парантропы
В 1948–1949 гг. Р. Брум находит нового парантропа крупнозубого – с
крупными зубами, костями и тяжелым черепом. Эта находка была самой примитивной
из всех найденных.
Долгое время шли споры о статусе австралопитеков – куда их
относить: к человеческой линии или человекообразным обезьянам? В 1950 году авторитетный
антрополог Ле Гро Кларк, детально изучив все находки и сравнив их с обезьянами,
пришел к выводу о том, что в зубной системе человека и обезьян насчитывается 12
различий: зубная система человека параболическая, а у обезьян зубы расположены
параллельными рядами; клыки обезьян имеют половые различия, и они из-за
большого размера при смыкании заходят друг за друга; у обезьян между верхними
резцами и клыками имеется диастема для выступающего снизу клыка и т. д.
Оказалось, что австралопитековые по отдельным признакам
обнаруживают сходство с человеком, а по другим – с
обезьянами, однако по большинству признаков они сходны с человеком. Ле Гро
Кларк (
С 1959 года в Восточной Африке велись раскопки супругами Мэри и
Луисом Лики. Первая находка была сделана в Олдувайском ущелье (Танзания) в
древнейшем слое. Это был гоминоид, названный «Зинджантропом бойсова» (Zinjanthropus boisei Leaky) (рис. 69, 70). «Зиндж» по-арабски означает
«Восточная», бойсова – в честь Чарльза Бойса,
финансировавшего исследования супругов Лики. В течение 30 лет работы супруги
Лики находили большое число окаменелостей животных, различные галечные орудия,
которые были названы «галечной культурой». Лики сам изготавливал подобные орудия и с их помощью за 20 мин мог
снять шкуру с антилопы. Однако Зинджантроп не мог быть творцом этой галечной
культуры по своей примитивности: его череп был с массивными гребнями, как у
гориллы, за огромные зубы его прозвали «щелкунчиком». Он был австралопитеком,
напоминавшим по своей массивности парантропа.

Рис. 69. Paranthropus boisei

Рис. 70. Череп Paranthropus boisei
В
результате дальнейших раскопок в Восточной Африке были найдены останки скелетов
австралопитеков, сосредоточенные в основном в области Восточно-африканского рифта
(рис. 71).
К
настоящему времени в Южной и Восточной Африке найдены и исследованы останки
нескольких сотен гоминид. Главная часть находок австралопитеков сделана на
африканском континенте, где они встречаются от Таунга на юге до Афары (Хадара)
на северо-востоке Африки.
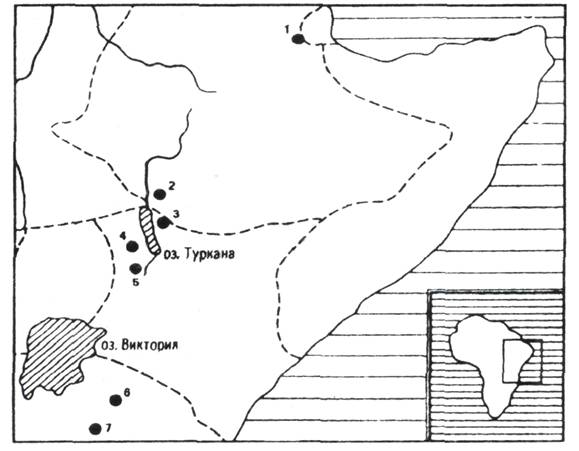
Рис. 71. Карта находок ранних
гоминид в Восточной Африке: 1 – Хадар; 2 – Омо; 3 – Кооби-фора; 4 – Лотагам; 5
– Канапои; 6 – Олдувай; 7 – Летоли
Основная
часть австралопитеков из Восточной Африки датируется периодом от 4,5–1 млн лет,
однако древнейшие двуногие гоминиды появились, вероятно, еще раньше. По
биомолекулярным данным, примерно 5,5–6,5 млн лет назад или еще раньше
«человеческая» линия эволюции отделилась от линии понгидов (шимпанзе).
Семейство гоминид характеризуется общим признаком – бипедией (хождение на двух ногах), которая связана с существенным
изменением условий внешней среды и образа жизни.
Особый интерес
представляет физический тип австралопитеков как переходная форма от понгидов к
человеку.
Прежде
всего это были двуногие формы с небольшими размерами мозга и крупными зубами,
покрытыми толстым слоем эмали.
Хорошо
известно, что все приматы обладают способностью поддерживать тело в
вертикальном положении, однако двуногое хождение человека коренным образом
отличается от прямохождения человекообразных обезьян. Основные изменения,
связанные с освоением бипедии гоминидами, сосредоточены в тазовом поясе и разгибательном
аппарате (рис. 72).
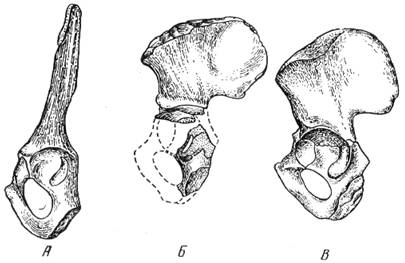
Рис. 72. Таз: А – шимпанзе; Б – австралопитека (Autralopithecus prometheus); В – современного человека
Важную
роль в преобразовании тазового пояса сыграли расширение подвздошной кости
вперед, усиление ее в средней части в области вертлужной впадины, укрепление
подвздошно-крестцового и тазобедренного сочленений, развитие связочного
аппарата, тазового пояса и крупных мышц – разгибателей бедра и пояснично-подвздошной
мышцы. Приматы, у которых отсутствует подобная структура, двигаются быстро,
наклонив туловище вперед. Что касается австралопитеков, то они имели хорошо
развитый человеческий разгибательный аппарат тазобедренного сочленения.
Применение рентгенографии и метода
компьютерной томографии подтвердило большое сходство во внутренней структуре
его таза и бедра и человеческого. Австралопитеки (грацильная группа, которая
дала начало гоминидной линии) отличались малым ростом (100–160 см), что
облегчало их двуногое хождение.
Головной мозг у австралопитеков с учетом массы их тела отличается
крупными размерами, его объем колеблется в среднем в пределах 440–450 см3 у
грацильных и 520–530 см3 – у массивных. Церебральный коэффициент,
определяемый по формуле Дюбуа (
E=kР 0,
56,
где E и Р – масса мозга и тела соответственно; k – коэффициент церебрализации
варьирует у австралопитеков в пределах 1,1–1,4, что в среднем в 2–2,5 раза
меньше, чем у современного человека. Этот показатель несколько выше по сравнению
с коэффициентом церебрализации современных высших человекообразных обезьян.
У австралопитеков особенно усложнились отделы головного мозга, в
частности нижняя теменная зона, связанная с ассоциативными функциями. На
мозговых слепках выделяются специфические области, характерные для человека и
связанные с процессами кинестезии и акустической коммуникации.
Череп австралопитеков небольшой долихоидного типа, свод черепа
выше, чем у шимпанзе.
По сравнению с понгидами в черепе австралопитеков обнаруживаются
прогрессивные признаки: у детеныша А. африканского надглазничный
валик отсутствует, затылочное отверстие сдвинуто вперед, размеры лицевого
черепа меньше мозгового. Лицевой отдел в целом уплощен, ноздри широкие и нижняя
челюсть развита слабо, а у парантропов она широкая и массивная.
Одновременно некоторые морфологические особенности австралопитеков
ставят их ближе к понгидам, чем к человеку: крупное лицо, выступающее вперед
(прогнатического типа), без подбородка, в зубной системе наблюдаются уменьшение
резцов и резкое увеличение моляров, покрытых толстым слоем эмали.
Анализ физического типа австралопитеков демонстрирует мозаичность
эволюции гоминид в миоценовую эпоху.
3.3.3.
Австралопитек афарский. Истоки рода человеческого
В 1973–1976 гг. в Северо-Восточной Африке (Эфиопия)
франко-американская экспедиция сделала ряд уникальных открытий. В 1974 г.
Дональд Джохансон в Афаре нашел почти полный скелет (40 %) костей,
принадлежавший женской особе ростом
Находка была настолько значимой, что вся экспедиция протанцевала
ночь напролет под песню «Битлз» «Люси в небе с бриллиантами». С того времени
находка получила свое название – «Люси», известное антропологам всего мира.
В последующие годы экспедиция Джохансона и Уайта нашла останки еще
30 индивидов того же вида, что и Люси. Анализ находок привел авторов к заключению,
что все представители вида обладают рядом примитивных признаков в зубной
системе, которые ставят их между обезьянами и человеком, т. е. они образуют
переходное звено между обезьянами и человеком, но с уклоном в сторону человека
(рис. 73).

Рис. 73. Д. Джохансон, М. Иди. Люси: истоки рода человеческого (
Емкость
черепа Люси составляла 380–450 см3 (рис. 73). Половой диморфизм
хорошо выражен.
Подобные
находки позже обнаружены в Танзании Ричардом и Мэри Лики. Все находки имели
одинаковые признаки в строении зубной системы, челюсть была V-образной формы, переходной от прямоугольной челюсти обезьян к
параболической у человека, сильное выступание лица (прогнатизм). В целом афарские
и танзанийские гоминиды, судя по скелету, были сильными существами, кости были
массивными с мыщелками и буграми для присоединения мощной мускулатуры. Четко
была освоена бипедия (рис. 74).
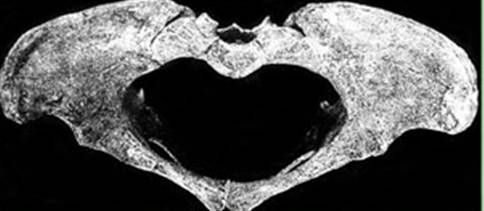
Рис. 74. Таз Люси,
демонстрирующий отчетливые признаки бипедии. Восстановлен из фрагментов О.
Ловджоем и Б. Браун
Найдены
следы стоп, отпечатанные в вулканическом пепле, окаменевшие с течением времени,
они были чисто человеческими. Руки были
длиннее, чем у человека, пальцы рук изогнуты, а запястные кости сходны с
обезьяньими, малый таз овальной формы, что явилось, вероятно, «дорогой платой»
за прямохождение. По мнению антропологов, такая форма таза затрудняла
роды, и определенная часть самок могла погибнуть во время родов (рис. 75).
Конечности
по пропорциям были сходны с плиопитеками и современными мартышкообразными.
Утолщение эмали на зубах свидетельствовало о питании семенами, которые требуют
усилия при их раскалывании.
Систематическое
название Люси – Австралопитек афарский
(Australopithecus afarensis).
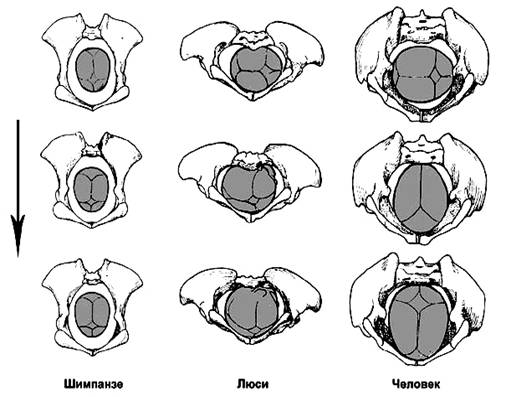
Рис. 75. Процесс родов и
бипедия – два фактора, определившие
строение таза современного
человека
Относительно
таксономии австралопитеков не все ясно. Высказано предположение, что
предшественниками ранних австралопитеков должны быть древнейшие
«преавстралопитекоидные», представители которых обнаружены в Кении (
В последние годы (1997–2002 гг.) в Африке обнаружен ряд
древних миоценовых гоминид, которые жили в период, когда произошла дивергенция
эволюционных линий шимпанзе и человека (6 млн лет назад). Систематическое
положение этих форм также остается спорным: сюда относят роды Сахелантроп,
Оррори и Артипитек.
Наиболее
древним гоминидом, найденным в пустыне севернее озера Чад около южного края
Сахары, является Сахелантроп, датируемый возрастом в 6–7 млн лет назад. В
2001 г. найден его череп в местечке Торос-Менелла (рис. 76).

Рис. 76. Сахелантроп (Sahelanthropus tchadensis) (
В
лицевой части черепа совмещены как весьма примитивные, так и относительно
прогрессивные черты: клыки слабые, размер мозга небольшой ~ 350 см3,
черепная коробка удлинена, что является характерным признаком для обезьян (рис.
76). Такая мозаика признаков свидетельствует о самых ранних этапах эволюции
гоминид. Кроме черепа, найдены фрагменты остатков еще пяти особей. В 2002 г.
международная группа ученых из 38 человек на основании находок описала род и
вид гоминид Sahelanthropus tchadensis. Находки показывают, что
древние гоминиды были расселены не только в Рифтовой долине Восточной и Южной
Африки, а распространены широко по всей Африке.
Оррори (Orrorin tugonensis) – другой древнейший гоминид, ходивший на двух ногах, обнаружен 25
октября 2000 г. при раскопках в Кении, назван человеком Миллениум.
Найдены кости около пяти особей в толще горных пород, возраст которых превышает
6 млн лет (рис. 77).

Рис. 77. Кости скелета
Orrorin tugenensis (
По
размерам напоминает современного шимпанзе. Судя по остаткам костей скелета,
данный вид проворно лазил по деревьям, совмещал также движения по Земле на
нижних конечностях. Питание растительное, но уменьшаются резцы, т. е.
эволюционная тенденция, согласующаяся с эволюцией человека.
В
1997–2000 гг. в долине Аваш (Эфиопия) найдены остатки скелета из миоценовой
эпохи (5,2–5,8 млн лет назад) А. ramidus (4,4 млн лет назад), в

Рис. 78. Остатки скелета
Ardipithecus kadabba

Рис. 79. Ardipithecus ramidus
Остатки
вида найдены там, где предположительно произрастали леса, но эти существа, судя
по скелету, могли быть двуногими (рис. 79).
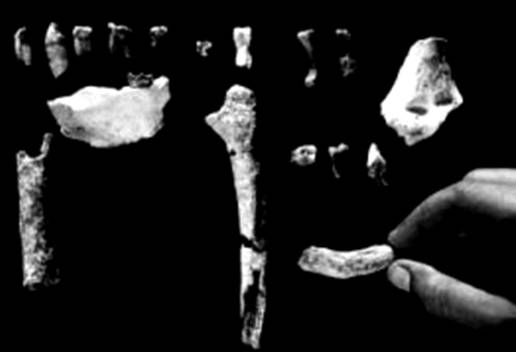
Рис. 80. Остатки скелета Ardipithecus ramidus
Этот факт истолковывается таким образом, что по господствующей теории
аридизация, изменение экологических условий послужили толчком для человеческой
эволюции, вытолкнув ранних гоминид на открытые саванны, где бипедия давала им
адаптивные преимущества. Однако подобная трактовка не согласуется с последними
находками.

Рис. 81. Australopithecus
anamensis
Australopithecus anamensis (Zeakey et. al., 1995) (рис. 81) появился в период между 4,17–4,12 млн лет назад;
предположительно по строению костей конечностей этот вид австралопитеков был
двуногим гоминидом. По строению зубов и челюстей он стоит ближе к поздним
ископаемым обезьянам (рис. 82).

Рис. 82. Останки
Australopithecus anamensis
По строению зубной системы вид занимает промежуточное положение
между Ardipithecus ramidus и Australopithecus afarensis. Авторы уверены в том, что A. anamensis
является предком A. afarensis.
3.4. Происхождение рода человеческого
Представители рода Homo впервые обнаружены 2 млн лет назад, до этого
на Земле жили только австралопитеки. Около 2,5 млн лет назад в эволюции
человека произошло важнейшее событие – дивергенция (кладогенез) с образованием
по меньшей мере двух линий гоминид: поздние массивные австралопитеки
(Зинджантроп бойсова, парантропы, австралопитек робустус и др.) и род Homo, первым достоверным представителем которого, по мнению
большинства исследователей, является H. habilis – «Человек умелый» (Лики, 1960) (рис. 83).

Рис. 83. Homo habilis
Развитие
массивных австралопитеков сопровождалось укрупнением размеров тела, усилением жевательного
аппарата, нарастанием физической мощи, замедлением роста черепа и энцефализацией. Массивные австралопитеки
считаются тупиковой линией, вымершей 1 млн лет назад. Причина их вымирания, по
мнению антропологов, состояла в их растительноядности. Отсутствие мясной пищи
не могло стимулировать охоту, орудийную деятельность, прогрессивное развитие
головного мозга. Таким образом, массивные формы австралопитеков были вытеснены
более грацильными формами гоминид.
3.4.1. Человек умелый – наиболее ранний представитель
рода
Homo
Предками
Homo считают афарских австралопитеков (Australopithecus afarensis),
относящихся к австралопитекам грацильного типа. Останки H. habilis обнаружены Л. Лики в 1960 г. в
Олдувайском ущелье в слое древностью около 2 млн лет. По фрагментам черепа
и посткраниального скелета он был схож с миниатюрными гоминидами, но с объемом
мозга большим, чем у зинджантропа (Australopithecus boisei) (1959 г.)
(рис. 84).
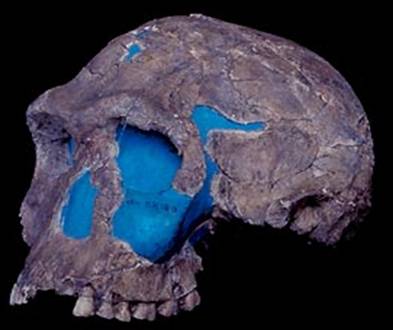
Рис. 84. Череп Homo habilis (экз. KNM УК
В
настоящее время исследованы ископаемые останки более 60 гоминид, найденных в
самых глубоких отложениях Олдувайского ущелья, которые в основном принадлежат
виду Homo habilis. Кроме того, недавно сделаны
находки черепов, зубов с переходными признаками между A. afarensis и H. habilis, что служит еще одним подтверждением схемы
филогенеза ранних гоминид, предложенной Джохансоном и Уайтом (
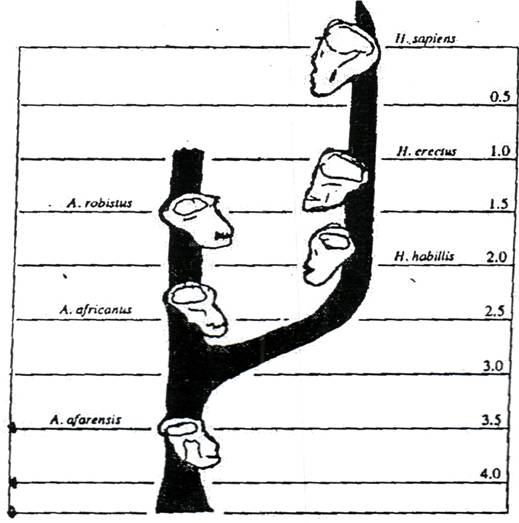
Рис. 85. Схема филогенеза
гоминид Джохансона–Уайта
(по вертикали млн лет)
В слое, где были найдены останки хабилиса (H. habilis),
обнаружены искусственно обработанные каменные орудия (отщепы). Они представляли
собой гальки небольших размеров с 5–6 сколами рабочего края, а противоположный,
необработанный конец закруглен и служил рукояткой. Такой тип орудия получил
название «чоппера» (анг. chopper), что означает ударник (рис.
86). Эта древнейшая культура человека получила название Олдувайской, или
«галечной».
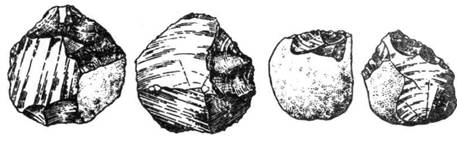
Рис. 86. Чоппер
До сих пор существуют спорные суждения о том, что предком Homo был
презинджантроп, однако по развитию мозга, этому важнейшему показателю, Homo habilis значительно превосходит
австралопитека. При равной массе тела объем черепа хабилиса составлял 645–660
см3, а у африканских австралопитеков – 450 см3, степень
церебрализации у хабилиса в 1,5 раза выше, чем у австралопитеков.
Высказано предположение (Tobias, 1983,
1987; Scheparts, 1993), что головной мозг хабилиса имеет
структурные основы для звуковой речи.
Отмечены также морфологические различия по сравнению с австралопитеками
в строении черепа и зубной системы (дентиции). Череп хабилиса расширен в
теменно-затылочной и заглазничной областях, размеры зубов уменьшаются, особенно
заклыковых моляров, что характерно для эволюции гоминидной линии.
Представляет особый интерес изучение кисти хабилиса. Структура
кисти сочетает в себе как прогрессивные, так и примитивные черты, в ней
сохранились признаки приспособления к древесному образу жизни. Привлекает
внимание расширение ногтевых фаланг, особенно на большом пальце. Этот
показатель является свидетельством формирования широких пальцевых подушек,
характерных для человека, развития кинестезического аппарата. Усиливается
силовой захват кисти, что было важно при изготовлении чопперов. Кроме
наследственного, расширение пальцевых подушек могло быть следствием
функциональной гипертрофии, которая наблюдается в практике длительного силового
захвата у представителей некоторых профессий современного человека. Такие
признаки вполне укладываются в представлении об эффективном использовании
хабилисом кисти при изготовлении примитивных каменных орудий.
Анализ кисти древнейшего афарского австралопитека показывает, что
на ней также наблюдаются понгидные и прогрессивные признаки: дистальная
укороченность пальцев, но без расширения конечных фаланг. Отсюда можно
заключить, что прогрессивные изменения кисти происходили довольно рано – еще у
плиплейстоценовых гоминид.
К роду хабилис относят и некоторых других представителей, найденных
у озера Рудольфа (нынешнее озеро Туркан) в Северной Кении. Здесь в 1968 г.
экспедицией под руководством Р. Лики открыты свыше 200 останков ископаемых
гоминид. Часть находок с восточного берега озера относится уже к роду Homo.
Особое внимание привлекает «череп 1470», найденный в 1972 г. в
нижних слоях отложений Кооби-Фора (восточное побережье озера Туркан) вместе с
другими остатками скелета. Реконструкция скелета показывает, что рост гоминида
составлял 156–160 см, емкость черепа – 752 см3, она несколько
превышает нижний предел емкости мозга питекантропа. По коэффициенту
церебрализации и анализу топографии сосудов эндокрана «череп 1470» сближается с
восточно-африканскими хабилисами из Танзании. Геологический возраст этого
гоминида по уточненным данным калий-аргонового метода составил 1,89–1,88 млн
лет назад.
К таксону Homo относят
также и некоторые другие находки из Эфиопии у озера Рудольфа (Homo rudolfensis)
из Южной Африки древностью 2–1,6 млн лет. Ископаемые материалы позволяют
заключить, что представители рода Homo жили в
Восточной Африке не позднее 2 млн лет назад (рис. 87, 88).

Рис. 87. Homo rudolfensis
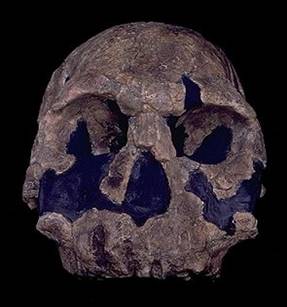
Рис. 88. Череп Homo rudolfensis
В связи с
тем, что хабилисы отличались высокой популяционной изменчивостью, некоторые
переходные формы, такие, как «череп 1470», таксономически не определены или
остаются предметом дискуссии.
Сенсационную
находку сделали грузинские археологи в 1991 г. под руководством Давида
Лордкипанидзе. Они описали посткраниальный скелет (кости конечностей и
позвонки) четырех примитивных представителей рода Homo, живших 1,77 млн лет
назад в районе города Дманиси (Грузия). В том же месте ранее обнаружены и
описаны четыре черепа. Впервые найдены кости древнейших людей, обитавших на
территории Грузии (рис. 89).
Грузинские
находки интересны тем, что в строении найденных черепов и посткраниальном
скелете сочетаются примитивные признаки, характерные для H. Habilis, и более
прогрессивные черты, сближающие их с H. erectus.
Анализ
материала грузинской находки позволил установить, что рост самого крупного из
четырех индивидуума составлял 145–166 см, вес – 40–50 кг. Эти данные вполне
сопоставимы с оценками, полученными ранее для H. habilis, а ранние
эректусы несколько крупнее (150,5–169 см при весе 45,5–70,6 кг).
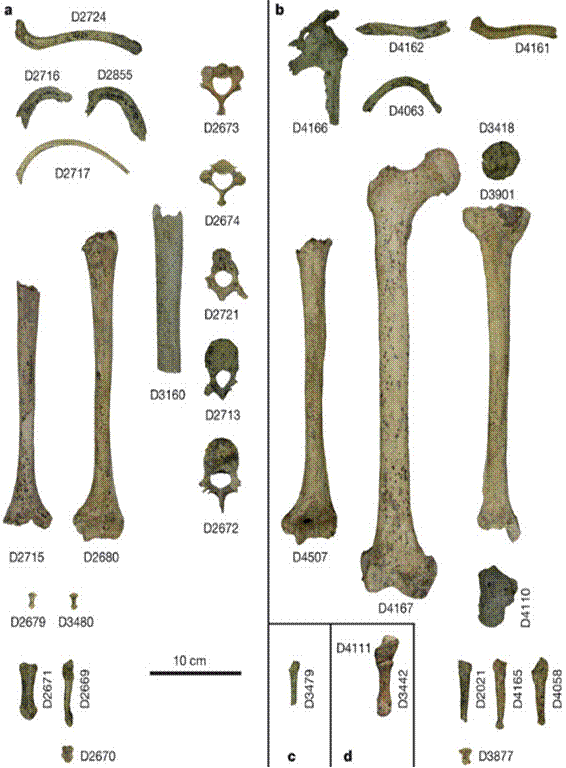
Рис. 89. Homo georgicus (2002 г.) 1,8 млн лет Грузия
Ноги
Дманисийских людей были почти такой же длины, как у эректусов и современных
людей, и длиннее, чем у австралопитеков. По-видимому, эти люди хорошо бегали и
могли преодолевать большие расстояния пешком. Это подтверждается и строением их
позвонков.
При
сходной с человеческой длине ног у людей из Дманиси руки длиннее и были похожи
на руки австралопитеков, особенно в строении плечевого пояса. По этому признаку
они напоминают хоббитов с острова Флорес. По коэффициенту церебрализации люди
из Дманиси стоят ближе к хабилисам, чем к эректусам. По строению позвоночника,
наоборот, они стоят ближе к эректусам.
Таким
образом, по строению скелета дманисийские люди занимают промежуточное положение
между хабилисами и эректусами.
Находка грузинских археологов заставляет пересмотреть
старые теории о миграции ископаемых людей за пределы Африки. Теперь ясно, что
первыми покинули Африку не эргастеры и эректусы, а именно переходные формы
между питекантропами и древнейшими представителями Homo, обладавшие небольшим
мозгом, как у хабилиса (David et. al., 2007).
Возможно,
питекантропы пришли на смену хабилисам приблизительно 1,6–1,5 млн лет назад.
В 1997 г.
Bermudes de Castro, Arsnaga и Gavbonel сообщили, что около миллиона лет назад
африканские популяции Homo erectus эволюционировали в новый вид Homo antecessor
(описанный ими же) (рис. 90). Они мигрировали в Европу, и в пещерах Испании
найдены их останки. По останкам скелета разных животных определили, что эти
люди были умелыми охотниками на крупных зверей. Геомагнитным методом определен
точный их возраст – около 800 тыс. лет. Они являются, таким образом,
самыми древними людьми, найденными в Европе. Голова этого гоминида обладала
мозаичностью черт неандертальца и современного человека, мозг – 1000 см3,
что дало основание авторам полагать, что вид был предком гейдельбержца, через
него неандертальца и сапиенса.
Высказано
и другое мнение, что сходство с гейдельбержцем и неандертальцем небольшое, а с
сапиенсами весьма значительное – в строении лицевой части черепа. Это позволяет
предположить, что предком современного человека является не H. erectus или H. ergaster, а H. antecessor.
Рис. 90. Череп Homo antecessor
3.4.2.
Древнейшие люди, или архантропы (Homo erectus)
Всех представителей прямоходящих людей объединяют под общим
названием архантропы, представленных одним видом Homo erectus.
Таксон H. erectus введен Кембеллом в 1962 г.
вместо Pithecanthropus erectus (Дюбуа,
1894) (рис. 91).

Рис. 91. Homo erectus
Архантропы
отличаются высоким надбровьем и мощным надглазничным валиком, низким сводом
черепа, объемом черепа 900–1100 см3, покатым лбом и плоским
затылочным отделом.
Первая находка человека прямоходящего, или питекантропа,
вызвавшая бурные споры и привлекшая внимание широкого круга антропологов,
принадлежит голландскому анатому и врачу Е. Дюбуа. Находка была сделана в
1891–1893 годы на
острове Ява. Это была первая находка человека прямоходящего по тем временам за
пределами Европы. В то время Э. Геккель предсказывал, что должно быть
«переходное звено» между обезьянами и человеком – питекантроп
(обезьяночеловек).
Отказавшись
от преподавательской деятельности, Е. Дюбуа устроился судовым врачом на
военном корабле и отправился на остров Суматру. Во время остановок корабля
Дюбуа занимался осмотром пещер, однако местные люди избегали пещер из-за злых
духов, и Дюбуа вел свои поиски по руслам рек, где можно было найти скелеты
животных.
В 1891
г. был найден третий верхний моляр, похожий по форме, длине и буграм на
человеческий, однако Дюбуа решил, что это обезьяний зуб. В 1892 году, продолжая
раскопки, он обнаружил в долине р. Соло крышку черепа в том месте (рис. 92),
где лежал зуб, а в

Рис. 92. Черепная крышка, найденная Э. Дюбуа ("питекантроп").
Homo erectus
Черепная
крышка занимала промежуточное положение между человеком и гоминоидом: лоб
покатый, надглазничные валики выделяются, как у обезьян. Емкость мозгового
черепа составляла 900 см3, на эндокране Дюбуа обнаружил отпечаток
извилины Брока, с которой связана речевая функция. Анализ черепной полости
показал, что она ближе к человеческой, чем к обезьяньей. Дюбуа дал своей
находке название Pithecanthropus и подарил Э. Геккелю с
надписью «Изобретателю питекантропа» (рис. 92).
Вернувшись
с острова Ява, Дюбуа предъявил находку ведущим ученым того времени: Кизсу,
Вудворту, Вирхову и другим. Она не была признана многими учеными как
«переходное звено». Вирхов считал, что останки принадлежат гиббону, а Кизс
полагал, что череп принадлежит человеческому дегенерату, которого ударили по
черепу, потому он стал плоским.
В 1895
г. на международном зоологическом конгрессе в Нидерландах 20 профессорам было
предложено высказаться за принадлежность находки Дюбуа человеку, «переходному звену» или обезьянам. Мнения разделились следующим
образом: бедренная кость большинством была признана человеческой, зубы и
черепная крышка были отнесены к переходному существу, были и такие,
которые посчитали, что бедренная кость и черепная крышка принадлежат разным особям.
После 10
лет безуспешной борьбы за признание находки Дюбуа спрятал ее от всех, а в конце
жизни решил, что нашел не «переходное звено», а останки гигантского гиббона. На
месте находки орудий труда обнаружено не было.
В 1936
г. геолог Р. Кенигсвальд продолжал поиски питекантропа в тех местах, где
работал Дюбуа на острове Ява.
В
местечке Моджекерто Кенигсвальд обнаружил грубо обработанные орудия с отщепами
– лезвиями. В период с 1936 по 1941 годы им было найдено 3 черепа и 3 нижние
челюсти. Один череп был детский, который сразу обратил на себя внимание
сходством с черепной крышкой, найденной Дюбуа в 1894 году: покатый лоб, низкий
свод черепа, мощные надглазничные валики и плоские височные кости.
Череп,
сходный с черепом обезьян, в то же время отличался большой емкостью в 900 см3,
что соответствует нижней границе вариации объема мозга современного человека.
Бедренная кость представляла резкий контраст по сравнению с черепной крышкой.
Эти противоречия породили новую волну дискуссий по находке Е. Дюбуа.
Кенигсвальд
добавил к своим находкам целый ряд фрагментов черепов (нижнюю челюсть с
премолярами и молярами, черепную крышку, затылочную кость и др.). Такое
сочетание примитивных черт в строении черепа с прогрессивными признаками
аппарата прямохождения соответствует в целом современным представлениям об
эволюции приматов. Известно, что аппарат прямохождения формировался гораздо
более длительное время (6–8 млн лет), а развитие головного мозга происходило
только в четвертичном периоде кайнозойской эры. Подтверждением служит тот факт,
что австралопитеки (Sahelanthropus tchadensis, Orrorin
tugensis, Ardipithecus kadabba и другие), жившие 6–8 млн лет назад и хорошо
овладевшие бипедией, имели примитивный головной мозг, близкий к обезьяньему.
Вероятно, на ранних этапах эволюции в связи с аридизацией климата и появлением
открытых пространств (саванн) освоение бипедии играло более важную роль в
процессах адаптации к изменяющимся условиям среды.
Находки
Кенигсвальда детально были исследованы антропологом Вейденрейхом (1873–1948
гг.), который отметил, что питекантропы, найденные им, несколько даже
прогрессивнее, чем находка Дюбуа. Один из черепов отличался большей массивностью, толщина на нижнем заднем
углу затылочной кости составляла
Синантроп и место H. erectus
в эволюции человека. Шведский
ученый Андерс в 1918 году в Китае около Пекина в каменоломне близ деревни
Чжокоудянь обнаружил кости ископаемых животных. Позже О. Зданский и В. Гранж на
тех же раскопках нашли 2 зуба, принадлежавших гоминиду. На основании этих
находок был выделен вид Sinanthropus pecinensis, а в 1929 г. Веньжун нашел и
череп синантропа (рис. 93).

Рис. 93.
Пекинский синантропа. Реконструкция черепа Ф. Вейденрейха
Анализом
фрагментов 44 особей обоего пола и
разного возраста занялся Ф. Вейденрейх. На
месте, где велись раскопки, было найдено множество каменных орудий и мощные толщи
пепла с остатками обгоревшего дерева. Находки относятся к среднему и началу
верхнего плейстоцена, т. е. межледниковому периоду. Орудия были сделаны из
песчаника, кварца и кремния: функциональное их назначение – отделение отщепов
для изготовления ручных рубил с двусторонней обивкой.
Кто был
предком H. Erectus, или переходной формой от H. habilis к H. erectus?
Чтобы ответить на этот вопрос, необходимо обратиться к
недавней находке представителя гоминид H. ergaster (рис. 94). Ранее его объединяли с африканскими
древнейшими людьми H. erectus, жившими 1,9–1,6 млн лет назад. Однако позже
большинство антропологов стали его выделять в отдельный вид по следующим
признакам: череп округлой формы, сильно развитые надбровные дуги, зубы мелкие,
особенно по сравнению с австралопитековыми. Отличается от эректусов более
тонкими костями черепа, слабым затылочным выступом. Объем мозга 880 см3.
Рис. 94. Череп Homo ergaster
Переход от
хабилиса к H. ergaster считается важнейшим качественным скачком в эволюции
гоминид.
H.
ergaster приписывают
два важнейших изобретения: обоюдоострые рубила, как зуб хищного зверя, и
использование огня (африканские кострища имеют давность более 1 млн лет).
Возможно, эти изобретения принадлежат их потомкам – эректусам.
Несомненно одно: при переходе от H. habilis к H. erectus произошли
два важных изменения: 1) резко увеличились размеры тела и 2) выросла доля
животной пищи в рационе, что тесно связанно с первым.
В 50-е годы XX в. Майр предположил, что все находки совместно с
теми, что обнаружены в Европе и Азии, относятся к одному и тому же широко
распространенному виду H. erectus.
Более поздние H. erectus, жившие 700–500 тыс. лет назад
имели головной мозг объемом около 1100 см3.
Полагают, что H. erectus произошел от H. ergaster примерно 1,6
млн лет назад и заселил юг Азии и Индонезийские острова.
Раньше полагали, что H. erectus вымерли около 300 тыс. лет
назад, однако по последним находкам на острове Ява возраст их определили в 50
тыс. лет, т. е. эректусы просуществовали очень долго и были современниками H.
sapiens.
Эректусы несомненно пользовались огнем и занимались каннибализмом:
найдены человеческие трубчатые кости, расколотые вдоль для извлечения костного мозга.
Характерный инструмент из камня H. erectus – это обоюдоострое
рубило, было универсальным орудием и в первую очередь для разделки туш.

Рис. 95. Нижняя челюсть Homo heidelbergensis
К числу важных находок относится нижняя челюсть
с зубами гоминида (рис. 95), найденная близ города Гейдельберга немецким
антропологом О. Шетензаком в 1907 г. Название гоминида определили по месту находки – гейдельбергский человек.
25 млн
лет 2
млн лет 12-14 млн
лет 4-5,5 млн
лет 1,5-1,9
млн лет 300
тыс. лет 40
тыс. лет Возраст
(млн лет)
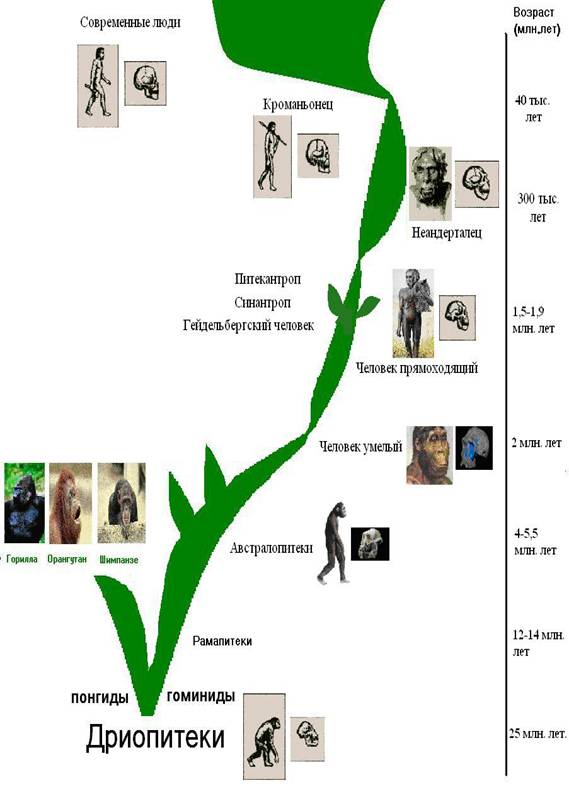
Схема 2. Эволюционное древо
Как
полагают некоторые ученые, подобные формы были переходными между
H. erectus и H. sapiens, жили примерно от 800 тыс. до 200 тыс. лет назад.
Нижняя челюсть была похожа на человеческую, без подбородочного выступа,
признака, который связывают с недоразвитостью или слабой развитостью речи.
Ранее их называли «архаичными H. sapiens». Согласно генетическому анализу линии
неандертальцев и современных людей разошлись 500–600 тыс. лет назад,
поэтому гейдельбержца нельзя считать «общим предком» тех и других, как это
следует из вышеприведенного эволюционного древа (схема 2). Гейдельбергский
человек является предком неандертальца, и это не вызывает сомнений (рис. 96).
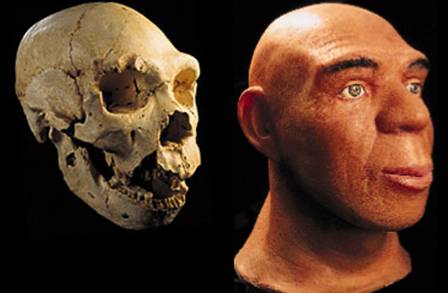
Рис. 96. Homo heidelbergensis
Как
видно на рис. 95, к особенностям строения, характеризующим нижнюю челюсть
гейдельбержца, относятся следующие: вырезка восходящей ветви слабо выражена,
суставной отросток и особенно его сочленованная часть наоборот хорошо выражены,
подобное строение нижней челюсти свидетельствует об ином характере жевательных
движений. По всем другим показателям челюсть гейдельбержца не отличается от
человеческой (клыки не конической формы, из зубного ряда не выделяются,
отсутствует диастема).
Близ
Гейдельберга в г. Шелль были обнаружены орудия, которые названы по месту их
находки – шелльские рубила.
Анализ
палеоантропологических материалов по древнейшему человеку (архантропам)
позволяет сделать некоторые обобщения.

Рис. 97. Неандертальцы (Homo neanderthalensis)
Человек
прямоходящий (H. erectus) имел широкий ареал расселения, который включал Яву,
Китай, Западную Европу, Южную и Восточную Африку. Бипедия была хорошо освоена,
аппарат локомации опережал в развитии головной мозг: в черепе, его лицевом
отделе и строении зубной системы сохранилось много обезьяньих черт. Имеются доказательства
деятельности по изготовлению орудий типа шелльских рубил.
Все они
жили в плейстоценовую эпоху и относятся к одному и тому же виду – Homo erectus.
Стоят ближе к современному человеку, чем австралопитековые.
Недавно
на индонезийском острове Флорес сделана находка скелета человека миниатюрных
размеров, позже обнаружены и другие скелеты подобного типа.
Крошечный
представитель Homo, прозванный исследователями «Хоббитом», жил всего 18 тыс. лет назад,
в то время, когда современные люди широко расселились по планете. Однако хоббит
напоминал уменьшенный вариант предков человека, которые были в сотни раз старше
и жили в другой части Азии.
Остров
располагается между Австралией и Азией и никогда не был соединен с Большой
землей, даже при низком уровне воды (во время отливов) расстояние между
материком и островом оставалось не менее
Однако в
50–60-е гг. XX в. археолог-любитель Т. Верховин нашел доказательства жизни
представителя современного человека на острове. Он нашел каменные орудия, возраст которых был
определен в 750 тыс. лет. Известно, что Homo
erectus (питекантропы)
не менее 1,5 млн лет назад жили недалеко от острова Явы, что позволило
Т. Верховину предположить, что некоторым из них удалось добраться до
Флореса по морю. Догадка Верховина подтвердилась. В сентябре 2003 г. достаточно
многочисленная экспедиция в составе индонезийских и австралийских
исследователей и 35 рабочих из местных жителей приступили к раскопкам в
долине Лианг-Буа, расположенной в западной части острова Флорес, и обнаружили
скелет взрослой женской особи.
Скелет
имел следующие особенности: скошенный лоб, сильно выступающие надглазничные
валики, мощная нижняя челюсть, череп, приплюснутый в области висков
(рис. 103). Он напоминал череп Homo erectus. Однако размеры тела были
необычными: масса тела составляла
В
середине 2004 года в Лианг-Буа на острове Флорес экспедиция обнаружила кости и
зубы, принадлежавшие еще шести другим особям, возраст которых от 98 до 13 тыс.
лет. Вид был назван Homo flоresiensis, предположительно он
представляет собой боковую ветвь эволюционного древа людей, его представителями
являются потомками изолированных островных популяций H. erectus (питекантропов).
Рис. 103. Homo floresiensis
Судя по каменным орудиям, питекантропы появились на острове 850
тыс. лет назад. В условиях островной изоляции они измельчали и сильно изменились,
в результате их потомки выделены в отдельный вид.
Рост этого вида людей на метр ниже H. еrectus, мозг, как у
шимпанзе, они были прямоходящими, лишенными волосяного покрова, пользовались
огнем, изготовляли достаточно совершенные каменные орудия и, вероятно,
охотились на крупных животных (стегодонов – карликовых слонов весом до
Местное население острова рассказывает много легенд о «маленьких
людях», живших вплоть до появления европейцев, которые освоили остров в начале
1500-х годов. H. flоresiensis
можно было бы отнести к «питекантропам – пигмеям». Однако карликовость этих
людей иного рода: она не связана с мутациями или нарушениями синтеза
соматотропина, как у настоящих пигмеев, у которых голова и мозг остаются
крупными.
По мнению авторов, их открытие меняет представление об адаптивных
возможностях рода Homo, можно ожидать в будущем находок ряда других видов и
популяций рода Homo, специализировавшихся и уклонившихся от общего
эволюционного древа людей (Morwood et. al., 2004).
3.4.3.
Древние люди, или палеоантропы
Палеоантропы (рaleos –
«древний», anthropos – «человек») – это
обобщенное название ископаемых людей, которых рассматривают как следующий этап
эволюции человека, следующий за архантропами и предшествующий неоантропам (рис.
97).
Палеоантропы жили в среднюю и позднюю плейстоценовую эпоху от
250–40 тыс. лет назад. Они представляли собой господствующую форму гоминид в
среднем палеолите в период их широкого расселения во внетропическом
пространстве: Крым, Северная Африка (Марокко), Передняя и Средняя Азия
(Узбекистан, пещера Тешик-Таш) (схемы 8, 9).
Первая находка палеоантропа сделана в Гибралтарской крепости в 1848 г., однако внимание ученых было
привлечено к ней в 1856 г., когда профессор Фульрот в долине реки Неандерталь
близ г. Дюссельдорфа (Германия) обнаружил останки человека, названного по месту
находки неандертальцем. Из пещеры был извлечен почти полный скелет: черепная
крышка, плечевые, локтевые, лучевые кости, ключица, таз, бедро.
Схема
плейстоцена и палеолита
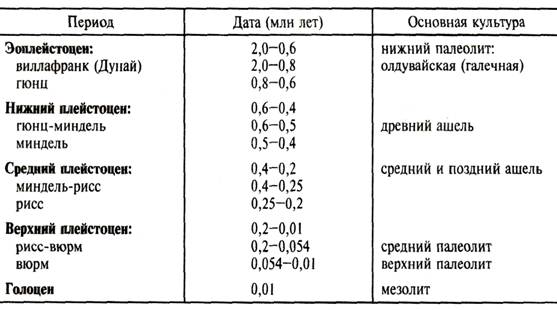
Древность
находки была признана не сразу. Рудольф Вирхов считал, что найденные останки
принадлежат субъекту с рахитическими отклонениями и серьезными деформациями в
черепе, может быть, даже идиоту. Однако Поль Брока и Томас Гексли признали
находку и установили древность в пределах 70–35 тыс. лет назад.
Схема
сравнительной стратиграфии
для
различных районов Евразии
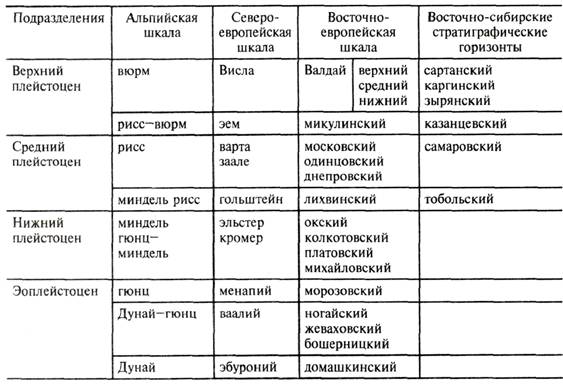
Неандертальцы
жили в приледниковых областях, поэтому хорошо были приспособлены к холодным
климатическим условиям. Они были небольшого роста (![]() ), что
ограничивает теплоотдачу, а сильно развитая мускулатура и мощный скелет
приспособлены к большим силовым и механическим нагрузкам.
), что
ограничивает теплоотдачу, а сильно развитая мускулатура и мощный скелет
приспособлены к большим силовым и механическим нагрузкам.
Мозг
неандертальцев крупный, несколько превышает даже объем мозга современного
человека. Череп долихоидного типа, с массивным надглазничным рельефом, свод
черепа низкий, лоб скошен. На затылке костный валик, называемый «шиньоном»,
служил, вероятно, для равновесия с лицевым черепом, который оставался
громоздким за счет прогнатизма верхней челюсти (рис. 98).
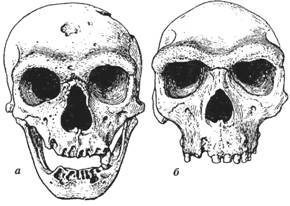
Рис. 98. Черепа
неандертальцев (по Broken-Hill, 1921): а
– из Chapelle-aus-Saints; б – из Rhodesia
У неандертальца
хорошо развита праворукость, «силовой захват», что привело к расширению
запястья и массивности кисти.
Палеоантропы
пользовались огнем, питались исключительно мясом, распространен был
каннибализм. У них появились первые мистические или религиозные верования,
хоронили своих умерших и украшали могилы цветами.
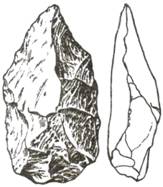
Рис. 99. Ашельское рубило
В период
Вюрмского оледенения в Европе была распространена основная культура палеолита –
мустье, названная по гроту Мустье – Ле Мустье во Франции. Мустье – это хорошо развитая каменная культура, отличающаяся
полифункциональностью, включавшая в себя десятки орудий постоянной формы.
Среди них особенно известны остроконечники, скребла, скобели, разнообразные отщепы, использовавшиеся при разделке туш (рис.
99). У неандертальцев имелись также деревянные копья для ближнего боя с
каменными наконечниками.
Отечественным
антропологом Г.А. Бонч-Осмоловским в Крыму около г. Симферополя в пещере Киик-Коба
(1924 г.), археологами А.П. Окладниковым в гроте Тешик-Таш (Средняя Азия,
Исходя
из анализа находок плейстоценовой эпохи и среднего и верхнего палеолита, можно
заключить следующее. Неандерталец (Homo sapiens neanderthalensis) является
более поздней формой архаичного сапиенса, произошедшей, вероятно, от
гейдельбергского человека.
Основанием
для такого вывода служат более 200 останков неандертальцев из Европы и Передней
Азии.
Большинство
находок соответствует первой половине вюрмского периода верхнего плейстоцена
(70–30 тыс. лет назад). Неандертальцы населяли приледниковые области,
были адаптированы к холодным климатическим условиям, сложился так называемый
арктический тип. Неандертальцы значительно продвинулись по культуре.
Мустьерская или Среднепалеолитическая культура (обоюдоострые рубила,
качественно заточенные по сравнению с рубилами Н. еrectus), отщепы
разнообразные, появились зачатки искусства (ожерелье из медвежьих когтей,
«скульптура», найденная недавно, духовность, религиозность).
Кроме
«классических» (рис. 100) в Западной Европе обнаружены и «атипичные»
неандертальцы древностью 250–200 тыс. лет. Находки сделаны в
Сванскомбе (Англия), Штенгейне (Германия) и пещерах Схул и Табун (Палестина).
Анализ
этих находок привел антропологов к гипотезе «пренеандертальца», т. е. к
происхождению современного человека от неандертальца. Была принята так
называемая стадиальная теория, по которой палеоантропы пришли на смену
архантропам. Допускалось смешение кровей (метисация) между неандертальцами и
современными людьми. Однако новые данные, указывающие на широкий размах
изменчивости в популяциях гоминид: мозаичность признаков, сочетание архаичных
черт с прогрессивными (объем мозга в 1350–1400 см3),
сопровождавшиеся массивностью затылочной кости с сильно выраженным наружным
рельефом, опровергли идею пресапиенса.

Рис. 100. Неандерталец
В настоящее время неандертальца не считают
самостоятельным видом, а выделяют как подвид таксона Homo sapiens – Homo
sapiens neanderthalensis
(Кэмбелл, 1966).
Неандертальцев
даже прогрессивного типа не относят к предкам современного человека, они
представляют боковую ветвь эволюции гоминид.
По
данным антропологов, в плейстоценовую эпоху в Европе существовал единый
эволюционный пласт, где происходило постепенное накопление сапиентных черт, но выделить
среди гоминид этой эпохи H. sapiens не удается. Наличие
межпопуляционных связей (метисация) позволяет допускать гипотезу об эволюции по
«сетевому типу», ослаблении или отсутствии эволюции по типу кладогенеза или
дивергенции.
Для
разграничения разных стадий эволюции гоминид антропологи используют типы
орудий, характерные для каждой стадии, например галечная культура архантропов
сменяется мустьерской неандертальцев.
В
заключение следует отметить, что происхождение человека от гоминоидных предков
подтверждается сходством их анатомии, физиологии, иммунологии, генетики и ряда
других показателей, общих для всех приматов.
Кроме
того, симиальная теория антропогенеза Ч. Дарвина основывается на находках
костных остатков ископаемых переходных существ (питекантропов и других
гоминидов), и поэтому в естествознании она в целом не вызывает сомнений. Однако
в дарвиновской гипотезе антропогенеза существуют некоторые противоречия и
загадки, не разрешенные до сих пор в связи с отсутствием прямых доказательств
или находок. В частности, в период с 5 до 4 млн лет назад не были обнаружены
останки гоминидов или они были крайне скудными (Хрисанфова, Перевозчиков,
2005).
Вызывает
сомнение вопрос о том, как могли сосуществовать столь разные формы антропоидов,
как австралопитеки грацильные (Australopithecus africanus), массивные
австралопитеки (Australopithecus robustus), еще более крупные зинжантропы
(Australopithecus boisei) и прогрессивные хабилисы в одной экологической нише.
Все они жили 2 млн лет назад. Не ясны причины потери волосяного покрова
человека, который защищал его по ночам от холода и сырости тропического леса.
В то же время сохраняется шапка волос на голове, ноздри повернуты вниз,
существуют значительные различия в зубной системе приматов, которые считаются
всеядными. Превращение питекантропа в современного человека за 4–5 тысячелетий
также считается маловероятным.
Костно-мышечная
система людей не приспособлена к древесному образу жизни, значит, для их
предков брахиация была невозможна. Что касается прямохождения, то оно миллионы
лет опережало в историческом плане изготовление орудий, тогда не понятно,
почему спустившись с деревьев, предки человека перешли к двуногому хождению.
Вероятно, труд только способствовал совершенствованию прямохождения, именно
прямохождение является предпосылкой освобождения передних конечностей для
труда. Укорочение и ослабление передних конечностей также трудно увязывается с
историческим развитием предков человека, так как сильные руки имеют гораздо
больше преимуществ в охоте и труде.
Зубы и
челюсти человека слабы для пережевывания сырого мяса, кишечник относительно
тела в 2 раза длиннее, чем у плотоядных, а у некоторых видов гоминид (например,
презинжантропа) челюсти значительно уменьшены, хотя огнем не пользовались.
Интересные
находки (1959–1980 гг.) были сделаны в районе водных источников (озеро Туркан и
река Омо). Австралопитеки и H. habilis жили в одно и то же время (2 млн лет
назад) и, вероятно, вели береговой образ жизни.
По
мнению автора теории Л.И. Ибраева (1985 г.), многие неувязки разрешаются, если
считать, что человек произошел не от лесных неуклюжих обезьян, а от береговых
обезьян (древнегреч. Pithecus najas naiad
– «нимфа»). Это были большеголовые, голые и двуногие прибрежные обезьяны. Они
обитали в неогене по берегам рек, ручьев, озер и др. пресноводных водоемов в
полусаванной предгорной области, занимались ловлей и собиранием моллюсков,
раков, черепах, грызунов, птичьих яиц, ягод, фруктов, корней растений,
насекомых и использовали для вскрытия раковин расколотую гальку, палки и кости.
Прежняя лесная жизнь позволила им приспособиться к береговому образу жизни. Они
имели пространственную координацию движений, хорошо развитые затылочную,
зрительную и теменную кинестетическую зоны коры головного мозга. Далее автор,
пользуясь следствиями антропогенеза, объясняет морфологические особенности
антропоидов и гоминидов, что позволяет разрешить многие противоречия,
изложенные выше.
Недавние
находки останков скелета гоминид в Дманиси (Грузия), на острове Флорес и
Испании заставляют пересмотреть теории миграции гоминид из Африки в Европу, а
также гипотезу антропогенеза в целом.
Учитывая
постоянно продолжающиеся поиски палеоантропологов и археологов по всему миру
остатков скелетов в отложениях третичного и четвертичного периодов кайнозойской
эры, возможны новые находки, которые позволят изменить существующие
представления и сформулировать новые теории антропогенеза.
3.4.4.
Современные люди, или неоантропы
Этап
эволюции и становление Homo sapiens sapiens по разным оценкам занял
длительный исторический период между 35–25 до 40–30 тыс. лет назад. Современный
взгляд на происхождение человека разумного допускает возможность сочетания
кладогенеза (дивергенции) с прямым эволюционным развитием отдельных линий
гоминид в полиморфных группах (Бунак, 1980). Вероятными предками сапиенса
называют гоминид, которые характеризовались сочетанием прогрессивных
(сапиентных) с архаичными чертами. Остатки скелета гоминида, найденного в
Вертешселлеше (Венгрия), имел объем мозга в 1300–1400 см3, как у
современного человека, в то же время затылочная кость была массивной и с
сильным наружным рельефом. Находки из Сванскомба (Южная Англия) и Штейнгейма
(Германия) древностью 250 тыс. лет дали основание по морфологическим
признакам выделить линию, ведущую к сапиенсу, и причислить их к таксону
пресапиенса.
Существует
также мнение о более позднем выделении линии сапиенса. Речь идет о
«неандертальской фазе», предложенной А. Хадличкой. Однако вопрос о роли
неандертальцев в происхождении человека остается дискуссионным. Если Н. erectus
рассматривается как обязательное звено происхождения человека, то
неандертальская фаза остается непризнанной в качестве таковой. Допускается
происхождение европейского сапиенса от пресапиенса или в результате поздней
миграции сапиенсов, возникших в Африке.
Сторонники
существования «неандертальской фазы» происхождения человека обосновывают свою
концепцию наличием между ними морфологической преемственности, однако
верхнепалеолитические сапиенсы Европы сходны не с классическими неандертальцами,
а с почти «сапиенсами» из Передней Азии (Кафзех-Схул), причем ранние сапиенсы
оказались на десятки лет древнее неандертальцев. Более детальные анализы
показали, что классические неандертальцы по морфологическим показателям черепа
не могут быть предком сапиенса.
В
современной антропологии доминирует мнение о том, что неандертальцев нужно
рассматривать как европейский вариант архаических сапиенсов и как боковую ветвь
эволюции человека в Европе. Неандертальцы не выдержали конкуренции с сапиенсами
и были вытеснены по всей территории Европы и Азии.
Обзор
основных гипотез позволяет заключить, что человек современного типа появился
около 100 тыс. лет назад. Между 40–35 тыс. лет назад H. sapiens широко расселяется по
Евразии и Африке и становится господствующим видом гоминид на Земле.
Человека
современного типа, или неоантропа, чаще называют кроманьонским человеком, хотя
самая первая его находка была сделана в 1923 г. в Уэльсе в пещере Павилад
(Англия). Скелет был окрашен в красный цвет из-за цвета почвы, где он пролежал,
и получил название «Красной леди». В 1852 г. из пещеры Ориньяк на Пиренеях
были извлечены 17 фрагментов скелета человека. В пещере были также обнаружены
орудия труда кроманьонца, которые получили название ориньякские (рис. 101).
В
1868–1894 гг. в гроте Кро-Маньон во Франции была открыта многослойная стоянка
древностью около 40 тыс. лет, обнаружены пять скелетов, принадлежащие
неоантропам. Подобные находки были сделаны в Белоруссии, на Украине, в России,
на Кавказе, на Памире, в Средней Азии и других районах. На этих стоянках при
раскопках находили орудия из морских раковин с проделанными в них отверстиями.
Такие же находки сделаны в гроте Гримальди, где обнаружено много ориньякских
орудий.

Рис. 101. Зарождение
«подлинно человеческой» духовной культуры:
Южная Африка, 72 000 лет
назад
Анализ
находок, проведенный П. Брока и Р. Верно, показал,
что все найденные скелеты принадлежат одному и тому же типу людей – кроманьонскому
человеку. Важнейшие особенности кроманьонца сводились к следующему: прямой лоб
без надглазничных валиков, высокий свод черепа, резко выраженное подбородочное
возвышение, без скошенных скуловых дуг. Для него характерны также череп с
широким лицом, развитые надбровные дуги, сильно выступающий узкий нос.
У
неоантропа височные и лобные доли мозга сильно развиты, что свидетельствует о
совершенствовании ассоциативных функций и корковых тормозных процессов.
Особенно хорошо развиты области головного мозга, связанные с речью. Известно,
что развитие лобных долей полушарий свидетельствует о высоком уровне процессов
коркового торможения. Клинические нарушения в префронтальной области полушарий
мозга сопровождаются возбуждением, агрессией, прожорливостью, половой распущенностью
и другими поведенческими актами, находящимися под волевым контролем. Отсутствие
контроля над поведением приматов привело бы к усилению агрессии в группах и их
взаимоуничтожению. Таким образом, лобные доли полушарий мозга послужили
морфологическим субстратом головного мозга, смягчающим агрессию и
обеспечивающим выживание человеческой популяции. Объем черепа доходит до 1800
см3, а размеры тела – до

Рис. 102. Реконструкция
кроманьонского человека (по Мак-Грегору)
В
отличие от неандертальца, для кроманьонца характерно удлинение предплечья и голени
соответственно по отношению к плечевой и бедренной костям.
Фундаментальные
изменения произошли в психической деятельности кроманьонца по сравнению с
другими гоминидами (неандертальцами и прямоходящими). Это отразилось на
социальности и орудийной деятельности: орудия становились разнообразными в
функциональном отношении и сложности изготовления. Развивались координация,
кооперация, альтруизм в деятельности первобытного общества. Появляются жилища,
сшитая одежда, музыкальные инструменты, зачатки искусства, наскальная живопись.
Анализ
находок кроманьонцев указывает на то, что с самого начала их существования
морфологические типы распадались на ряд расовых вариантов. В частности, в
черепах взрослой женщины и юношей, найденных в гроте Гримальди (Италия), обнаружены
резко отличительные признаки по сравнению с
кроманьонцами, найденными в том же гроте: мозаичное сочетание расовых черт,
совмещающих в себе европеоидные и негроидные черты, – выступание челюсти
(прогнатизм) и слабо выступающий широкий нос. Такое же сочетание расовых черт
обнаружено на черепах кроманьонцев древних захоронений Воронежской области
(30–25 тыс. лет назад), в скелете мужчины отмечены наряду с
европеоидным признаком (выступание носа) и негроидный признак (прогнатизм и
широкий нос).
3.5. Прародина человека и гипотезы моно-
и полицентризма
Прародина
человека является предметом дискуссий до настоящего времени. Различные
представители гоминид были широко распространены на трех континентах: в Африке,
Азии и Европе, что позволяет допустить возможность возникновения человека на
любом из них. Однако с учетом данных палеоантропологии, археологии и
молекулярно-генетических исследований более вероятной прародиной человека могла
быть Африка. Такого мнения придерживался и Ч. Дарвин. Эволюция приматов
происходила в плейстоценовую эпоху под влиянием целого ряда
экологических и геологических факторов. Все приматы – обитатели тропиков,
т. е. гоминиды по своему происхождению являются тропическими организмами,
которые уже позже в связи с изменением условий существования приспосабливались
к разнообразным климатическим условиям и экотопам.
Наиболее
значительными событиями в эволюции гоминид являлись бипедия (двуногое хождение)
и орудийная деятельность. Оба события обоснованы с точки зрения тропической
экологии и истории геологии. Тропики характеризуются постоянной высокой
температурой в течение круглого года и четкой сменой двух сезонов – сухого (без
дождей) и дождливого. Приспособление к высокой температуре у млекопитающих, в
том числе приматов, связано с наличием шерстного покрова, который служит
хорошим изолирующим материалом. Человек же потерял волосяной покров, и
терморегуляция при этом происходит за счет учащения дыхания и интенсивности
потоотделения. В связи с этим гоминиды вынуждены были держаться около крупных
водных источников: реки Омо, озера Туркан и Олдувая. Не случайно большинство
находок гоминид сделаны в Олдувайском ущелье, у озера Туркан.
Что
касается бипедии, то она явилась одним из морфологических механизмов,
понижающих площадь поверхности тела, подвергающейся воздействию прямых
солнечных лучей. Кроме того, прямохождение имеет гораздо больший энергетический
эффект по сравнению с хождением на четырех ногах как по скорости, так и по
выносливости. Человек превосходит по выносливости таких четвероногих, как
зебра, антилопа и других, африканские охотники могут загнать свою четвероногую
жертву до коллапса.
В
результате вулканической активности и горообразовательных процессов леса были
вытеснены биотопами саванн, и возникла мозаичная экосистема с изолированными
биотопами. Одновременное усиление радиационной активности (в Южной Африке
добываются урановые руды) стимулировало мутационные процессы, и создались
условия для формирования переходных форм и эволюции гоминид. В тропиках в
настоящее время выделяются следующие биотопы: 1) тропический лес с 2–5 ярусами
растительности с лианами; 2) широколиственные тропические леса; 3) колючий лес
с открытыми пространствами и крупными животными; 4) саванна – тропический
биотоп с травоядными животными и хищниками; 5) пустыни и полупустыни; 6)
горные ландшафты.
Дело в
том, что именно в Южной и Восточной Африке были широко расселены
австралопитековые, человек умелый и человек прямоходящий. Существуют две точки
зрения по поводу места происхождения человека: моно- и полицентризм.
Сторонники
концепции моноцентризма полагают, что современный человек появился в одном
центре с расселением на другие территории («миграционная гипотеза»). Возможным
вариантом моноцентризма (П. Эндрюи, Л. Шотт и другие) могло быть возникновение H. sapiens в Африке с миграцией в Европу. По мнению
других сторонников моноцентризма, человек возник в Азии и, отделившись от
азиатских гоминид, мигрировал в Европу (рис. 104).
Рис. 104. Монофилетическая
гипотеза, или гипотеза африканского происхождения человека. Человек
современного типа возник в Африке и распространился оттуда на все континенты,
вытесняя других
представителей
рода Homo (Stringer, 1989)
Гипотеза моноцентризма подтверждается современными
молекулярно-генетическими исследованиями, которые доказывают генетическую
гетерогенность населения Африки, что может быть следствием высокой мутационной
активности. Исходным центром распространения разнообразия населения могла быть
Восточная Африка.
Гипотеза полицентризма, выдвинутая американским антропологом Ф.
Вейденрейхом (1943 г.), предполагает, что H. sapiens мог возникнуть в 4
центрах: 1) в Юго-Восточной Азии – развитие шло от питекантропа к австралоидной
расе; 2) в Восточной Африке, где синантроп дал начало монголоидной расе и
американским индейцам; 3) Южная Африка стала
центром возникновения негроидной расы; 4) Передняя Азия – центр европеоидной
расы от палестинского гоминида, хотя следы миграции из Передней Азии в Европу
не известны.
В отечественной антропологии разработана также гипотеза «широкого
моноцентризма» (Рогинский, 1947). Предполагается происхождение современного
человека на обширной территории Средиземья, охватывающей Восточную Африку,
Переднюю Азию и Юго-Восточную Европу. Этот процесс сопровождался смешением
кровей (метисацией) между разными группами гоминид, появлением переходных форм,
предоставляющих обширный материал для отбора и формирования рас, хотя
существуют доказательства, что всем расам дал начало современный человек (Homo sapiens).
Существует также и современный вариант полицентризма – так
называемый мультирегиональная гипотеза.
По этой гипотезе, Н. еrectus из Африки расселился в другие регионы, и
современный человек (H. sapiens) возникает в нескольких центрах соответствующих
территорий, где сосредоточены современные расы (рис. 105).
Рис. 105. Мультирегиональная гипотеза. Человек современного анатомического
типа возник независимо на разных континентах из разных ветвей Homo erectus.
Единообразие людей поддерживалось системой миграций (Stringer, 1989)
При анализе генетического материала по митохондриальной ДНК (мхДНК)
выявлен большой разброс по датировкам, что свидетельствует о том, что предок
современного человека, судя по данным мхДНК («митохондриальной Еве»), мог быть
скорее эректусом, чем сапиенсом, что хорошо согласуется с гипотезой
полицентризма.
Несмотря на различие взглядов ученых по поводу места и времени
происхождения человека, моно- и полицентристы, согласно концепции моногении,
сходятся во мнении, что у современного человека предок был один, и все расы
представляют собой один вид – Homo sapiens.
3.6.
Основные
этапы происхождения человеческого
общества. Социогенез
Возникновение и эволюция человека тесно
связаны с проблемой развития общественных отношений, или социогенезом. На начальных этапах антропогенеза важнейшим механизмом
становления гоминид были биологические факторы: мутагенез, борьба за
существование и естественный отбор. Однако общественный труд являлся
направляющей силой естественного отбора, который избирательно «подхватывал»
преимущественно те изменения в биологической организации гоминид, которые в
дальнейшем способствовали развитию трудовой деятельности и общественных
отношений.
В целом соотношения между генетическими и социальными факторами на
различных этапах эволюции являются наиболее сложными, т. к. сам человек по
своей природе является биосоциальным феноменом. Трудно себе представить
возникновение сознания, речи с точки зрения генотипа человека, так как такие
чисто человеческие качества не сводятся только к генетическим факторам, хотя
они создают биологическую основу развития мышления, речи и языка. Для их
развития необходима социальная среда. Известно, что у детей, изолированных от
общества с самого рождения, речь и прямохождение не развиваются.
По мере социализации человека происходило также постепенное
уменьшение формообразующей роли естественного отбора. Завершением антропогенеза
явилось возникновение Homo
sapiens sapiens, существование которого опосредовано через социальную среду.
Сообщества животных характеризуются наличием определенных условий,
способствующих их упорядочению и укреплению: сложные системы связи,
специализация, различие функций, стабильность состава (Дюсберг, 1981).
Человеческое общество состоит из групп,
коллективов, классов, наций и государств, и все социальные категории активно поддерживаются
всеми членами общества. С самого начала возникновения человеческого общества
главной силой его укрепления выступают производственные отношения, и в связи с
этим следует считать, что начало первобытного человеческого общества совпадает
с изготовлением орудий труда.
Реконструкция начальных этапов социогенеза
представляется наиболее сложной из-за отсутствия прямых сведений. Поэтому
восстановление исторической картины ранних этапов развития общества возможно
гипотетически, исходя из данных палеоантропологии, археологии раннего палеолита
и этологии современных приматов, наиболее близких к предкам человека.
Модельными
сообществами могут служить стада павианов и макак с хорошо организованной
иерархической структурой, а также шимпанзе, который генеалогически наиболее
близок к человеку (рис. 106).
Следует подчеркнуть, однако, что до сих пор исходного предка
человека не удается идентифицировать с каким-либо из ныне существующих
понгидных видов. Здесь может представлять интерес для сравнения с гоминидами групповое
поведение понгид, которые, по наблюдениям этологов, обладали определенным
интеллектуальным потенциалом для перехода к орудийной деятельности (Бутовская,
1994).
В эволюции гоминид важную роль могла играть также способность
приматов к обучению.
Афарские австралопитеки являются самыми
ранними гоминидами, они жили, по мнению антропологов, бродячими группами по
20–30 особей на обширной территории; судя по морфологическим показателям, они
сохраняли связь с лесной средой. Австралопитеки не имели постоянных убежищ,
лагерей и баз, их иногда называют «первобытным человеческим стадом».
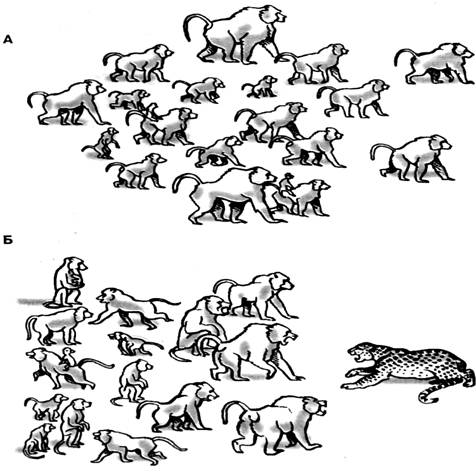
Рис. 106. Стадо павианов на марше (А) и при возникновении опасности (Б)
Плотность
популяции австралопитеков олдувайской эпохи оценивают в 0,001–2,5 индивида/км2
в долине реки Омо. Считают, что продолжительность жизни австралопитеков не
превышала 20 лет в среднем, а прирост населения за 1000 лет составлял 10–20 %.
Австралопитек
африканский использовал в пищу кроме растений
и мясо. Об этом свидетельствовали найденные Дартом в пещере Макапансгат (Южная
Африка) «кухонные кучи».
Основой
жизни австралопитеков могли быть собирательство и охота. Орудия
«костно-зубо-роговой культуры» австралопитеков использовались не только для
охоты, но и для защиты от хищников. В первобытном стаде складывалось устойчивое
объединение со сложившимися иерархическими отношениями. По мнению французского
антрополога А. Валяда, в связи с тем, что самки погибали во время родов, в
группах самцов всегда было больше, что вело к конфликтам из-за самок. Афарские
австралопитеки заплатили дорогую «цену» за прямохождение, так как организм
перестраивался, овальной формы таз не был достаточно укреплен. В Южной Африке в
пещерах были обнаружены 17 скелетов австралопитековых со следами насилия,
причем установлено, что удары наносились длинными трубчатыми костями. Однако
Р. Дарт считал, что в группах австралопитеков убийства невозможны, а
обнаруживаемые повреждения на черепах имели посмертный характер с целью
вскрытия черепа и извлечения мозга для поедания.
Однако
нельзя исключать внутристадные столкновения, которые служили фактором
дестабилизации стадных отношений. Механизмом снятия агрессии между самцами в
стаде служило удлинение эстрального цикла самок. Ожесточенные драки могли
возникать также при распределении пищи. По наблюдениям Р. Телеки, подобные
драки вспыхивали между самцами шимпанзе при распределении мяса. Американская
исследовательница Д. Гудалл, наоборот, утверждает, что шимпанзе охотно делится
добычей с другими особями. О дележе добычи в прямом смысле слова в данном
случае можно говорить с осторожностью, так как кусок мяса, урываемый тайком,
нельзя называть дележом пищи.
По
другим наблюдениям, доступ к добыче получают без ограничения доминантные особи,
а субдоминанты выпрашивают пищу у особей высокого ранга или тайком пытаются
откусить кусок.
Австралопитеки,
в отличие от шимпанзе, обладая зубо-костно-роговой культурой, могли добывать
больше мяса. Тем более что у австралопитеков была развита кооперация, и они
охотились на более крупную дичь. В связи с разным иерархическим статусом не все
члены сообщества получали равную долю мяса.
Что
касается орудийной деятельности, то не вызывает сомнения, что австралопитеки могли
раскалывать камни с целью выбора пригодных для использования обломков, однако
целенаправленная орудийная деятельность у них на уровне всей популяции
появилась, вероятно, позже.
Полагают,
что австралопитеки африканские могли иметь стоянки для хранения орудий, которые
служили также центрами жизнедеятельности, куда самцы могли приносить добычу
(Дерягина, 1994).
Дальнейшее
развитие социальности гоминид связывают с появлением хабилисов (H. habilis). В первом слое Олдувайской долины обнаружены
искусственные сооружения из базальных блоков, уложенных в виде круга. Их
называют «обитаемыми горизонтами». Обнаружено около 20 таких «домов-баз»,
представляющих собой скопления костей, каменных орудий, возможно, они служили
охотничьими стоянками. Есть предположение, что олдувайские гоминиды не только
подбирали остатки туш – жертв крупных хищников, но и сами выступали охотниками
на крупных травоядных животных. Обнаружены трещины от ударов на черепах
антилоп, найдены кости животных с насечками от каменных орудий.
В большей
части гоминиды Олдувая, вероятно, занимались собирательством и подбором падали.
Однако охота играла важную роль для дальнейшего развития социальных отношений в
популяции гоминид. Охоту на крупных животных, особенно на тех, которые ведут
стадный образ жизни, трудно себе представить без загонного метода. Поскольку
вооружение хабилисов было несовершенным, то необходимо было отбить животного от
стада и шумом, криком, камнями, огнем и другими способами загнать его к ущелью
или крутому обрыву: животное падало с обрыва и разбивалось, а человеку
оставалось его добыть.
Таким
образом, охота являлась эффективной формой трудовой деятельности, которая
стимулировала кооперацию, организованность, объединяла членов группы в трудовом
процессе и демонстрировала преимущества коллективизма.
Разделение
труда имело место уже у олдувайских хабилисов между самцами и самками. Первые
преимущественно занимались охотой, а вторые – собирательством. Разделение
функций также явилось фактором развития социальности: мужские особи занимались,
возможно, транспортировкой охотничьей добычи в стойбище самкам и детям, а также
для дележа. Такое проявление альтруизма в популяции могло быть предметом
естественного отбора. Происхождение альтруизма – достаточно сложный вопрос и
его проявление и толкование также неоднозначны. Считается, что некоторые формы
альтруизма (например, реципрокный или взаимный альтруизм) могут быть
результатом отбора (Красилов, 1986); в то же время постоянно возрастающие сроки
детства в эволюции, необходимость кооперации в охоте и групповой
жизнедеятельности, забота о стариках как носителях опыта поколений, и другие
факторы, вероятно, понижали остроту и роль борьбы за существование.
Разделение
труда привело к взаимозависимости между самками и самцами. Самки и дети
нуждались в мясе, которое добывалось самцами, а самцы, наоборот, нуждались в
растительной пище, собираемой самками.
У
человека умелого могли возникнуть зачатки семейной жизни, т. е. самец
обеспечивал пищей не всех особей в группе, а только одну самку, однако это не
могло быть строгой необходимостью, так как самки и дети могли кормиться
самостоятельно вместе с другими самками без самцов, такой способ жизни
сохранился у многих первобытных народов, доживших до наших дней.
Однако,
по мнению исследователей, семья могла возникнуть в составе более крупного
многосамцового объединения, характерного для приматов вообще. Кроме того, самец
в одиночку не мог бы защищаться от хищников и охотиться на крупную дичь.
Переход
от первичного человеческого стада к первобытным социальным отношениям
обозначить трудно, так как отсутствуют четкие социальные и морфологические
критерии; по мнению многих исследователей, он занял большой исторический период
между средним и верхним палеолитом (0,2–2 млн лет).
Антропологи
связывают ранние этапы социогенеза с орудийной деятельностью гоминид.
Исторически достоверный этап начала трудовой деятельности отражен в анализе
находок олдувайской культуры Homo habilis.
По
свидетельству антропологов, питекантропы вели более подвижный образ жизни,
связанный с миграциями, охотой и освоением новых территорий. Их стойбища
использовались более интенсивно по сравнению с хабилисом. Кроме более или менее
постоянных стойбищ обнаружены стоянки, где разделывались туши животных. Судя по
большому количеству изготовленных орудий, питекантропы в стойбищах
задерживались долго, т. е. проявляли сезонную оседлость. Обнаружены также
«мастерские» по производству орудий. Строились примитивные «жилища» с полами,
вымощенными галькой, хижины имели длину 7–15 и ширину – 4–6 м. Очаг располагался
в центре хижины. Охота как основной вид деятельности питекантропов
подтверждается большим количеством костей разных животных, в т. ч. и слонов,
найденных на стойбищах (стоянках).
Этнографические
данные о численности групп охотников, размеры жилищ и интенсивность их
использования позволяют восстановить уровень социальной организации эректусов.
Ученые считают, что группы питекантропов составляли 40–60 особей, в т. ч. 20
взрослых человек.
Что
касается взаимоотношений, то многие исследователи склоняются к мнению, что в
группах питекантропов нередко возникали кровавые столкновения. Как показал
анализ повреждений на скелетах питекантропов, проведенный американским
антропологом Ф. Вейденрейхом, они имеют прижизненное происхождение и нанесены
каменными орудиями типа ножей и топоров, хотя драки из-за самок также не
исключаются.
Существует
мнение, что с началом изготовления орудий в первобытном человеческом стаде
усилились конфликты, драки стали частыми и более ожесточенными, тем не менее
первобытные коллективы отличались достаточной устойчивостью. Если бы не было
механизмов стабилизации коллектива, то первобытное общество не развилось бы до
высоких форм социализации. Однако следует подчеркнуть, что в первобытном
человеческом стаде шла постоянная борьба между эгоистическими и
стадно-коллективистическими формами поведения. Без сомнения, первые вытеснялись
вторыми, иначе стадная организация не переросла бы в более высокие категории
социальности.
Вероятно,
на самых ранних этапах антропогенеза в истории первобытного общества животный
эгоизм подавлялся, отдавая предпочтение добыванию и распределению пищи, половой
жизни. Этот процесс усиливался действием естественного отбора. Выживали
коллективы, в которых социальные связи (альтруизм, кооперация, коммуникация и
другие) хорошо выражены и находятся под контролем отбора.
Следующим
уровнем социализации, вероятно, было изготовление более совершенных ручных
рубил из камня. На этом этапе у орудий, кроме рабочего края, появилась
определенная форма, что свидетельствует о повышении уровня абстракции. Ученые
отмечают связь между функциональной активностью (сноровкой) руки и развитием
определенных структур мозга. У палеоантропов, в частности, отмечается развитие
ассоциативных зон в теменной дольке (области надкраевой и угловой извилин). Однако
на современных приматах установлено отсутствие соответствия изменений макро- и
микроструктур мозга техническим приемам, используемым гоминидами.
На этапе
неандертальцев социогенез развивался в сторону повышения активности охоты,
сплочения коллективов и дальнейшего совершенствования орудий труда. Гоминиды
специализировались в выборе объекта охоты. Палеоантропы чаще селились в пещерах
и специализировались в охоте на пещерного медведя.
Во
Франции в пещере Ля Бони обнаружено жилище размером 5 ×
По
данным археологии и палеонтологии, древние люди убивали отдельных членов своего
коллектива, возможно, ударами, нанесенными каменными орудиями, иногда кости
людей обожжены, как у животных, употребляемых в пищу. На скелете (ребрах и
других костях) обнаружены раны в стадии заживления после прижизненных травм от
каменных орудий.
Однако
существует много доказательств альтруистического поведения древних людей. В
частности, обнаружены останки неандертальцев с деформированными суставами,
вероятно, за ними был уход, иначе они не дожили бы до глубокой старости.
Охота
была одним из главных источников жизни, объединявших гоминид в сообщество; пища
распределялась и тем, кто не участвовал в охоте: женщинам, детям и больным.
Охота велась на крупных животных, хорошо заметных в высокой траве (носорогов,
слонов). Для добывания таких крупных обитателей у неандертальцев не было
соответствующих орудий. Поэтому более вероятным методом могла быть загонная
охота: загоняли крупную дичь в топкие места, убивали камнями. Судя по размерам
площадок для «разделки» туш, численность охотников могла быть 10–35 человек.
Сложная загонная охота и последующее распределение добычи требовали от древних
людей способности к планированию и кооперации. Кроме того, без системы
коммуникации и наличия членораздельной речи трудно представить сложное
общественное поведение гоминид.
Олдувайские
орудия отличались примитивностью: отмечается небольшой процент орудий по
сравнению с числом необработанных камней. Число ударов при изготовлении
каменных орудий хабилисом было 3–7, а при изготовлении ашельских чопперов
количество ударов доходило до 50–80.
По мере
социализации человека формообразующая функция естественного отбора постепенно
ослабевает, и 40 тыс. лет назад он полностью прекратил свое действие на
развитие человека. Генетическая программа Homo sapiens как вида сформировалась полностью,
и с тех пор у человека разумного никаких изменений в наследственном аппарате не
происходило. В то же время большое значение приобретало формирование социальной
личности через воспитание и обучение. При незначительных морфологических изменениях
человек сделал гигантский скачок в психическом развитии. Культурная эволюция
имеет колоссальную скорость распространения по сравнению с генетической.
На
определенном уровне развития общественных отношений мог возникнуть родовой
строй. По мнению ученых, это могло произойти в период между средним и верхним
палеолитом. Именно к верхнему палеолиту происходит завершение формирования
сапиенса как вида; трудовая деятельность и культура в верхнем палеолите
поднимается на новый уровень развития: археологи обнаруживают разнообразие
конструкций жилищ, сделаны находки целых стоянок – поселений, «мастерских».
Население этих жилищ могло перерасти в производственные организации типа рода,
общины; появилось многообразие орудий труда из различных материалов, приделывались
рукоятки, наконечники, значительно повышающие силовой эффект. Одновременно
появляются различные виды искусств: пещерная живопись, мелкая пластика
(статуэтки), музыкальные инструменты (рис. 107–109).
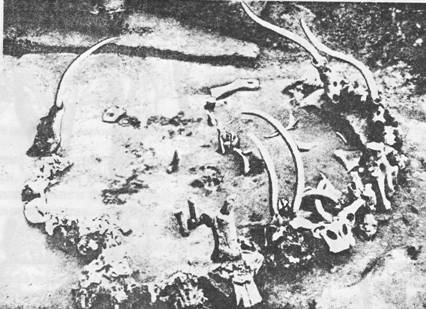
Рис. 107. Мезинские жилища
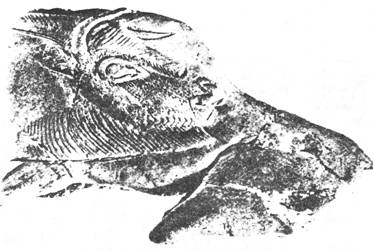
Рис. 108. Скульптура бизона

Рис. 109. Статуэтки,
относящиеся к эпохе верхнего палеолита (цит. по М.Д. Гводовер)
Преимущество
в выживании могли иметь те коллективы, в которых легче преодолевалось
противоречие между кооперативными действиями всех членов (изготовление орудий
труда, сложные формы охоты и др.) и возможностью присвоения результатов труда
(Рогинский, 1947).
На
начальных этапах становления родового строя коллективы разбивались и
перемешивались с другими группами в связи с введением экзогамии. Экзогамия
состоит в запрете половых отношений между членами одного и того же рода и
разрешении их между разными сообществами, племенами и группами.
По
мнению исследователей, первой исторической формой общественного регулирования
отношений между полами мог быть один из видов экзогамии – дуально-родовой
групповой брак. Дуально-родовой брак ставил запрет половых отношений с
сородичами, что, вероятно, предотвращало, хотя и неосознанно, нежелательные
генетические последствия
близкородственных браков, кроме того, в результате гетерозиса повышалась
жизнеспособность потомства. Взаимобрачие между двумя определенными родами называется
направленной экзогамией. Такая древняя форма брака заключала также супружескую
общность, которая состоит в том, что мужчины одного рода считаются мужьями всех
женщин другого рода. Древность дуально-родовой экзогамии подтверждается
этнографическими данными. Реконструкция подобного вида брачных отношений
осуществляется исходя из анализа сохранившихся брачных институтов у некоторых
племен. Например, в Австралии живут племена, которые разделены на две половины
и внутри них брачные связи запрещены, в то же время мужчины одной половины
племени с самого рождения считаются мужьями всех женщин второй половины и
наоборот.
«Парадокс
человека», по мнению Тейяра де Шардена, заключается в сравнительной ничтожности
морфологических изменений по отношению к огромному скачку в психической сфере.
Социализация
древнейших гоминид происходила на основе биологических и генетических
предпосылок. Афарские австралопитеки из Южной Африки и антропоморфные обезьяны
жили в сходных условиях лесосаванны. По образу жизни, поведению и психике они,
вероятно, также были сходными, что подтверждается исследованием эндокранов
ископаемых останков. Исходя из этого, естественно предположить, что для
реконструкции социогенеза исходными моделями могут служить приматы с наиболее
хорошо сложившейся групповой системой, которые экологически и морфологически не
были строго специализированы. В таких группах должны были гармонично сочетаться
элементы группового поведения – как агрессивного, так и дружелюбного. В
качестве примера можно обратиться к групповой системе макак, шимпанзе, горилл,
павианов и др. Для всех них характерны большие мультисамцовые группы.
Можно
полагать, что в древнейших группах человека было по несколько женщин и мужчин.
Отдельные
исследователи считают, что группы древнейших людей напоминали сообщества
павианов гамадрилов, которые представляют собой гаремные объединения. Однако
наблюдения показывают, что гаремы возникают в ответ на неблагоприятные условия
среды. Павианы гамадрилы живут на скалистых участках и в случае засухи, отсутствия
корма сообщества распадаются на односамцовые группы. Гаремы не могли быть
основой организации первобытных людей, так как самцу, чтобы удерживать гарем,
нужно постоянно быть внимательным и заботиться о том, чтобы снять напряжение в
гареме. Гаремы неэффективны также и в плане организации охоты, так как самцы не
могут оставлять самок без присмотра, чтобы их не разобрали молодые самцы.
В
гаремах поведение особей лишено лабильности и гибкости, оно более стереотипное
и не располагает к развитию.
Социогенез
неразрывно связан с проблемами понижения уровня агрессии в сообществах,
выработки механизмов или буферов агрессивного поведения, которые позволяют не
только тормозить, но и предотвращать агрессию. Преодоление конфликтов и
кровопролитий в группах достигается не ужесточением
иерархии в группах, а наоборот, развитие кооперации и альтруизма позволяет
понижать уровень агрессии.
Против
агрессивности в группах приматов существуют и другие механизмы. Особо агрессивные
особи обезьянами изгоняются из сообщества, а самки для спаривания выбирают
менее агрессивных самцов.
Развитие
альтруизма как одного из фундаментальных свойств человека имеет особенно важное
значение для социальной организации. Альтруизм является генетически
недетерминированным свойством человека, как считают социобиологи, а
социокультурное поведение, наследуемое через воспитание, традиции, искусство,
приобретается экстрагенетическими способами.
1. Антропогенез, приматология, морфофункциональная характеристика
приматов.
2. Низшие приматы. Семейство полуобезьян, морфофункциональные
особенности долгопят и лемуров.
3. Характеристика широконосых обезьян: капуцинообразные и
цебусовые.
4. Классификация и характерные особенности узконосых обезьян:
мартышкообразных и человекообразных (Hominoidea).
5. Морфологические особенности, образ жизни и среда обитания
мартышкообразных обезьян.
6. Высшие узконосые, или человекообразные обезьяны: гиббоновые,
понгиды (орангутаны, горилла и шимпанзе), их морфологическая характеристика и
среда обитания. Социальная жизнь: взаимоотношения высших обезьян в группах,
иерархия.
7. Характеристика гоминид.
8. Морфологическая и функциональная характеристика современного
человека.
9. Геологические и экологические факторы, приведшие к возникновению
примитивных приматов в конце мелового периода мезозойской эры.
10. Геохронология кайнозойской эры (международная шкала
подразделений по Дж. Симпсону).
11. Три периода «радиации» в эволюции приматов.
12. Суть «тарзиоидной гипотезы» происхождения приматов в
соответствии с филетической линией: плезиадапиды → омомиды →
антропоиды.
13. Пути миграции приматов в Америку: гипотезы «восточного пути» и
«западного пути». Автохтонный путь развития американских широконосых обезьян.
14. О месте дриопитеков в эволюции приматов, «проконсулоидная
гипотеза» происхождения высших приматов.
15.
Восстановление филогенетических линий по данным биомолекулярных часов.
16.
Ископаемые формы австралопитеков и их морфологические особенности.
17.
Австралопитеки как охотники: доказательства и значение в эволюции.
18. «Австралопитек прометеев» и «зубо-костно-роговая
культура».
19.
«Парантропы», их морфологические особенности.
20.
Место австралопитеков в эволюции приматов.
21.
Аппарат прямохождения, его становление, трансформация тазового пояса, связочной
и мышечной систем.
22.
Коэффициент церебрализации австралопитеков.
23.
Морфологическая характеристика черепов австралопитеков.
24.
Австралопитек афарский (Люси) – родоначальник человечества. Морфологические
особенности скелета, связанные с прямохождением. Признаки, характеризующие Люси
как переходное существо от обезьяны к человеку.
25.
Таксономическое положение австралопитеков. Характеристика находок из Сахары,
Кении, Эфиопии.
26.
Важнейшее событие, произошедшее около 2,5 млн лет назад, положившее начало двум
линиям гоминид.
27.
Назовите ископаемый гоминид, который был автором каменных орудий, найденный
Луисом Лики.
28. Морфологическая
характеристика H. habilis, место находки его останков и значение в эволюции
гоминид.
29.
Характеристика Олдувайской галечной культуры.
30.
Морфологические особенности черепа и кисти H. habilis. Другие представители
рода H. habilis, «череп 1470», его эволюционное значение.
31. Суть
острых дискуссий по находке H. erectus, их причина.
32.
Находка Е. Дюбуа, ее характеристика. Причины ее непринятия зоологическим
конгрессом.
33.
Находки В. Кенигсвальда.
34.
Находка синантропа, его морфологическая характеристика.
35.
Предполагаемый предок H. Erectus, или переходная форма от H. habilis к H.
erectus.
36. H.
ergaster и его морфологическая характеристика.
37.
Находка нижней челюсти гейдельбергского человека, его морфологическая
характеристика.
38.
Морфологическая характеристика архантропов, их систематическое положение. Место
эректусов в эволюции человека.
39.
Неандертальцы, их ареал расселения и морфологическая характеристика.
40.
Мустьерская культура. «Классические» и «атипические» неандертальцы.
41. Гипотезы
происхождения Homo sapiens по «сетевому типу» и ослабления типа кладогенеза в
плейстоценовой эпохе.
42.
Сенсационная находка грузинских археологов и новый подход в теории
антропогенеза в связи с этой находкой.
43.
Палеонтологическая связь находки в районе города Дманиси (Грузия) с открытием
человека на острове Флорес.
44.
Место H. antecessor, найденного в Испании, на эволюционном древе людей.
45.
Длительность периода эволюции и становления сапиенса.
46.
Гоминиды, которые по морфологическим и функциональным признакам могли быть
предками Homo sapiens.
47. Идея
выделения линии, ведущей к сапиенсу, или таксона «пресапиенса».
48.
Гипотеза «неандертальской фазы» эволюции человека.
49.
Исторические периоды становления человека современного типа.
50.
Время и место находки скелета «Красной леди».
51.
Ориньякские орудия, принадлежавшие кроманьонцу, их характеристика.
52.
Особенности черепа кроманьонца.
53.
Морфологические изменения, произошедшие в строении головного мозга неоантропа.
54.
Фундаментальные изменения, произошедшие в психической деятельности кроманьонца
по сравнению с неандертальцами и эректусами.
55.
Морфология H. flоresiensis. Гипотеза сходства с H. erectus.
56.
Влияние островной изоляции на морфологию находки на острове Флорес.
57. Материалы
палеоантропологии, геологии, археологии и молекулярно-генетических
исследований, позволяющие считать прародиной человека Африку.
58. На
основании чего принято считать, что тропическая экология и история геологии
могли способствовать развитию орудийной деятельности и бипедии у гоминидов?
59.
Причины того, что древние гоминиды держались около крупных водных источников и
там же чаще обнаруживаются их останки.
60.
Геологические факторы, которые могли способствовать образованию новых биотопов,
изолировавших группы гоминид и усиливавших их эволюцию.
61.
Место происхождения гоминид: теория моноцентризма, или «миграционная» теория.
62.
Автор и положения гипотезы полицентризма.
63.
Гипотеза «широкого моноцентризма» Я.Я. Рогинского.
64. Суть
мультирегиональной гипотезы полицентризма.
65.
Значение охоты на крупных животных (загонная охота) в развитии общественных
отношений в группах ранних гоминид.
66.
Орудийная деятельность гоминид и развитие социальности.
67.
Причины агрессивного поведения гоминид и механизмы снижения агрессии.
68.
Значение экзагамии в развитии родового строя.
69.
Дуально-родовой брак. Направленная экзогамия.
70.
Развитие социогенеза у неандертальцев.
71.
Соотношение генетического и социального на этапах эволюции гоминид.
72.
Связь между социализацией гоминид и снижением формообразующей роли
естественного отбора.
73.
Производственные отношения, связанные с орудийной деятельностью гоминид.
74.
Ранговая иерархия у ранних гоминид и распределение пищи в группах.
75.
Развитие социогенеза у хабилисов: дома – базы, «обитаемые горизонты», их
значение в развитии социальности.
76.
Элементы разделения труда у хабилисов между самками и самцами.
77.
Начальные этапы кооперации у гоминид. Альтруизм.
78. Суть
формулы Тейяр де Шардена: «Парадокс человека» заключается в ничтожности
морфологических изменений по отношению к огромному скачку в психической сфере.
79. Роль
гаремных объединений в эволюции приматов.