Раздел 2. Организованность
биосферы и ее усложнение с эволюцией жизни и
биокосных систем планеты
Тема 6. Эволюционные изменения
интегральных характеристик биосферы: изменение общей биомассы и продуктивности,
изменение энергетики биосферы,
изменение информационного «фонда» и эволюция биологического круговорота
1. Изменение общей биомассы и продуктивности
2. Изменение энергетики
биосферы
3. Изменение
информационного «фонда».
4. Эволюция биологического круговорота.
По Э.И. Колчинскому (1990), к числу параметров, характеризующих
биосферу как целостную систему, следует отнести ее общую биомассу и
биологическую продуктивность, энергетику, информационный «фонд» и
общепланетарный биологический круговорот. Проблема их эволюции поставлена еще в
трудах В.И.Вернадского, но из-за своей сложности и отсутствия надежных данных
далека от решения.
1.Изменение
общей биомассы и продуктивности
К вопросу об изменении
биомассы биосферы В.И.Вернадский
подходил неоднозначно. И в настоящее время существуют сторонники идей как
постоянства биомассы, так и увеличения ее количества в ходе эволюции жизни.
Большинство биологов считает, что в ходе истории живых организмов возрастало
общее количество живого вещества, о чем свидетельствовали нарастающее
разнообразие организмов, завоевание все новых экологических зон и
дифференциация экологических ниш. Геологи, биогеохимики и философы скептически
относятся к возможности однозначно решить этот вопрос. Так, В.И.Виноградов (1975) пришел к выводу,
что ни видовой состав биосферы, ни ее пространственное строение не определяют
количества живого вещества. Биомасса биосферы всецело обусловлена наличием тех
элементов и определенных условий в пределах земной коры, которые необходимы
для построения органического вещества. Ученый полагал, что уже первичные
организмы в относительно кратчайший срок (геологического времени) использовали
все потенциальные возможности для увеличения биомассы.
Решение проблемы зависит,
прежде всего, от возможности количественной оценки биомассы в современной
биосфере и биосферах прошлого. В.А.Успенский
(1956) исходя из постоянства коэффициента фоссилизации органического углерода
(0,8 % органического углерода уходит в захоронение с осадочными породами) и
показателей его содержания в осадочных породах попытался определить суммарную
биомассу биосферы за все время ее существования, взятое условно за 600 млн.
лет. Даже при столь заниженном возрасте оказалось, что годовая продукция органического
углерода в современной биосфере превышает объем средней годовой продукции в
биосферах прошлого в 29,5 раз. Полученный результат хорошо согласуется с
данными о содержании углерода в осадочных породах палеозоя, мезозоя и кайнозоя,
которые показывали постепенное возрастание содержания органического углерода в
суммарной биомассе фоссилизирующего углерода. Так, среднее содержание углерода
в палеозойских отложениях — 0,62%, в мезозойских — 0,68% и в кайнозойских —
1,32%. Другими авторами также были произведены расчеты по динамике содержания
органического углерода в породах разного возраста.
А.Б.Ронов
(1964) пришел к выводу, что факт увеличения содержания органического углерода
в более молодых породах объясняется, прежде всего, увеличением биомассы и
биоактивности растений, усилением перехода их остатков в ископаемое состояние,
а также потерями летучих соединений древними отложениями. Н.М.Страхов (1971) и Н.Б.Вассевич
(1973) отмечают, что неизвестно, какая часть отмершей части переходит в
ископаемое состояние. Так, в эволюционном ряду хвойные леса — лиственные леса
— травянистые сообщества шло уменьшение процента органики, поступающей в
захоронение. Фитоценозы последних этапов развития биосферы отличаются все
более замкнутым биологическим круговоротом, почвами с все более быстро
мигрирующей органикой и с все меньшим содержанием органических веществ,
выключенных из биологического круговорота.
Кроме величины биомассы в
эволюции жизни и биосферы существенную роль сыграло изменение продуктивности
экосистем. Биомасса большинства травянистых растений почти полностью
минерализуется в 1—2 года, в то время как биомасса стволов, корней, ветвей
деревьев остается живой десятки и сотни лет. Поэтому аккумулятивное значение
травянистого покрова в биосфере не меньше, а, возможно, больше, чем лесной
растительности. Особенно это касается таких явлений, как гумусообразование и
биогеохимия углерода, азота, кремния, кальция, калия, фосфора. По подсчетам В.А.Успенского (1956), в результате
биологических процессов в ненарушенных экосистемах (дыхание, разложение,
жизнедеятельность беспозвоночных, грибов, бактерий и др.) продуцируется
огромное количество углекислоты 13,5×1010 т/год. Интенсивность
циклических процессов образования живого вещества за время существования
биосферы, по мнению В.А.Успенского, возросла в 29 раз. Травянистые растения
сыграли в этом не последнюю роль.
В ходе эволюции биосферы
менялось соотношение между продуцентами, консументами и редуцентами. Если в
океане, древнейшей части биосферы, биомасса продуцентов составляет всего 1/16
часть, то на суше биомасса растений более чем в 99 раз превышает биомассу
животных и микроорганизмов.
Большинство биологов считает,
что живое вещество биосферы, используя непрерывность поступления космической
энергии, развивалось по принципу «самоуправляемого расширенного воспроизводства»
биомассы, численности организмов, их разнообразия и растущей сложности.
2.Изменение энергетики биосферы
К.А.Тимирязев рассматривал
хлорофилл в качестве посредника между рассеянной солнечной энергией и энергией
синтезируемых в растениях органических веществ. В. Анри подчеркивал, что жизнь препятствует превращению полезной
энергии в теплоту, и был уверен, что в ходе прогрессивной эволюции в биосфере
увеличивается количество энергии, аккумулированной организмами. В.И.Вернадский в 20 – 40-х гг. XX в. подсчитал количество энергии,
связанной с живым веществом, — n×1019 ккал.
Биосфера как автотрофная
система оказалась в состоянии не только обеспечивать космической энергией свои
потребности, но и аккумулировать значительные ресурсы энергии. Увеличение
биомассы и накопление ископаемого органического вещества изменяют энергетику
биосферы. Только в месторождениях горючих ископаемых сконцентрировано более
1×1013 т органического вещества (А. И.Перельман, 1973), а
всего в осадочных породах – около 3,48×1015 т (Н.Б.Вассевич,
1976). Энергия, накопленная в осадочных породах, равняется 1×1025
Дж (Р. С. Волскис, 1982).
По В.А. Ковде (1985), гумусовая оболочка суши содержит примерно такое
же количество связанной энергии, как и надземная биомасса. Ископаемые
источники энергии (уголь, сланец, торф, нефть, газ), созданные биосферами
прошлого, на 2 – 3 порядка выше. При этом не учитывается энергия, связанная в
рассеянном органическом веществе толщ осадочных пород.
Огромное количество энергии накопилось
в земной коре и в почве за всю историю биосферы. Кроме того, значительная часть
энергии, аккумулированная ранее живым веществом, теперь представлена в форме
поверхностной энергии дисперсных частиц в зоне выветривания, а также
расходуется во многих геологических процессах. Живое вещество, участвуя в
раздроблении горных пород на тонкодисперсные коллоидные вещества, передает им
дополнительную энергию.
В целом энергетические
ресурсы биосферы складываются из притока солнечной энергии (в год) – n×1020-21 ккал, учтенных
запасов ископаемой энергии – n×1022-23, энергии биомассы планеты — n×1020, энергии биомассы
суши – n×1019,
энергии гумусовой оболочки почв и мелководий – n×1019-20,
энергии, связываемой ежегодно фотосинтезом, - n×1017-18, энергии, связываемой
ежегодно в фитомассе земледелия, - n×1012-13 ккал.
Чем больше суммарная
поверхность, тем больше энергия. Наибольшей поверхностной энергией обладают
глины, илы, ископаемые угли, чернозем, которые способны сорбировать из окружающей
среды различные соединения. Аккумуляторами энергии являются и сульфиды: чтобы
восстановить серу (S6+→ S2-) и железо (Fe3+→ Fe2+), надо затратить энергию, и она в скрытом виде содержится
в пирите и других сульфидах. При окислении на земной поверхности сульфиды
отдают энергию окружающей среде. Поэтому в зоне окисления сульфидных руд
повышается температура. Во многих странах на участках разработки сульфидных
месторождений руд известны «колчедановые пожары». В данном примере энергия
выделяется и в химической форме, так как при окислении пирита образуется серная
кислота, разрушающая окружающие породы.
Благодаря энергетической
функции биосфера оказалась способной поддерживать жизнь на планете, умножать
численность и усложнять виды организмов, обеспечивать потребность гетеротрофных
организмов — животных и человека в энергии.
По Г.Ф.Хильми, в поверхностной оболочке Земли постоянно накапливается
превратимая энергия, что обеспечивается благодаря выработке у растительного
покрова адаптации к максимальному захвату солнечной энергии. На континентах
оно достигается путем увеличения фотосинтезирующей поверхности (образо![]() вание листовой мозаики, непрерывный растительный покров, его
ярусная структура, дифференциация растений по сезонам вегетации).
вание листовой мозаики, непрерывный растительный покров, его
ярусная структура, дифференциация растений по сезонам вегетации).
В целом в биосфере
используется лишь 0,1 — 0,2 % годовой величины солнечной радиации,
аккумулированной фотосинтезом. На суше она колеблется в пределах 0,3 — 0,46 %
(оптимум 1 — 2 %), эффективность использования энергии фитопланктоном ниже
(0,04% энергии, достигающей поверхности океана). Обнаружено увеличение
энергетической эффективности экосистем различного эволюционного возраста (по
отношению продукции к биомассе). В ландшафтах хвойных лесов, возникших в
середине пермского периода, отношение продукции к биомассе составляет 0,54 —
0,55, в ландшафтах с покрытосеменными деревьями, возникших в середине мела, —
0,59 — 0,68. В ландшафтах с травянистыми покрытосеменными (степи, саваны),
возникших в неогене, достигает 0,96. По мнению А.И.Перельмана, возрастание ежегодной продукции на единицу биомассы
следует рассматривать как показатель прогрессивного развития экосистем. В.Ф.Левченко считает, что эффективность
наземных экосистем по использованию солнечной энергии возросла за последние
540 млн. лет в 100 раз.
Энергетическая эволюция
биосферы связана с возрастанием целесообразности живых систем: рост общей
экономичности обмена веществ и круговорота биомассы, уменьшения непроизводительных
потерь при большей работоспособности, более высокой надежности при большей
активности, нарастании тенденции к автоматизации (И. И.Шмальгаузен, 1968),
усилению гомеостатических механизмов и др. Для биоэнергетики клетки высших растений
характерен высокий консерватизм, по-видимому, уже на начальных стадиях жизни
была выработана весьма совершенная структура внутриклеточных энергетических
процессов.
Одним из наиболее общих и
наглядных способов характеристики энергетической эффективности биосферы, по К.М.Завадскому, может стать критерий
средней выживаемости особи у доминирующих в биосфере таксонов. Повышение средней
выживаемости особи стало главной стратегией в прогрессивной эволюции живого. В
то же время повышалась энергетическая эффективность живого вещества в биосфере.
Если у низших организмов, представителей древних биосфер, средняя выживаемость
особи очень мала — 1/109 из числа родившихся, то у высших она
достигает 10 — 30 %. Таким образом, значительно возрастает вероятность
выполнения видовой функции воспроизведения; у низших организмов способ
воспроизведения неэкономичен, а у высших сократились непроизводительные
расходы биомассы. Это обеспечило захват жизнью новых экологических ниш, интенсификацию
энергетических процессов в биосфере, повышение миграции элементов и др.
Появление гомойотермных животных
было подлинной революцией в энергетике биосферы, так как расходование ими громадного
количества энергии для поддержания физиологического оптимума потребовало
дополнительных запасов биомассы на низших уровнях трофических цепей.
Высокая эффективность
использования энергии на уровне элементарных физиологических процессов
обеспечивает выживание организмов. Сложившаяся структура использования энергии
на разных уровнях организации живого является фактором, гарантирующим
стабильность биосферы. В ходе ее эволюции была создана такая структура жизни,
при которой распределение энергии по разным уровням и эффективность ее
использования на каждом уровне обеспечивали целостность и устойчивость
биосферы.
Мы находимся лишь в начале
пути познания энергетических закономерностей эволюции биосферы как целостной
системы. Пока приходится ограничиваться положением о том, что по мере прогрессивного
развития жизни усиливается накопление в биосфере превращаемой энергии и
обогащение этой энергией других поверхностных оболочек Земли.
3.Изменение информационного «фонда»
В современной биологии и
экологии под информацией, как правило, понимают меру многообразия и сложности
организации жизни и биосферы в целом. В ходе истории живых организмов
наблюдалось увеличение информационной «емкости», рост количества форм
организмов. Изменение информации — это главная тенденция и в эволюции биосферы,
которая выражалась в возрастании ее многообразия и структурированности: увеличение
количества геохимических барьеров, рост дифференцированности
физико-географического и геохимического строения биосферы и т.д.
Первым кибернетический подход
к эволюции живого применил М. М.
Камшилов (1974). Он пришел к выводу о неизбежности ускорения роста
информации в биосфере по мере возрастания ее сложности. Для количественной
оценки роста информации применил данные о видовом разнообразии организмов. В
отдельные периоды широкого распространения зон с теплым и влажным климатом
(карбон, мезозой) объем живого вещества был больше, чем в современной биосфере.
Однако разнообразие современных физико-географических условий (наличие аридных
и гумидных зон, высокогорий и т.д.), в которых обитают организмы, обусловливает
большее разнообразие видов животных и растений или, иными словами, большее
количество биологической информации.
По В. А. Кордюму (1983), в биосфере находится громадное количество
генетической информации, которая благодаря различным способам неполовой
передачи (симбиоз, трансформация, трансдукция) постоянно воздействует на
организмы. Это позволяет организмам быстро приобрести новые признаки.
Предположив широкий обмен генетической информации между очень отдаленными
таксонами, ученый признает реальной единицей эволюции не вид, а биосферу в
целом.
По теории симбиогенеза
профессора биологии Амхерстского университета Л. Маргулиса (L. Margulis, 1970), в основе эволюции эукариот находится механизм включения
генетического материала микроорганизмов в наследственные клетки растений или
животных. Возникающие в результате новые генетические системы — гибриды
бактериальной и растительной или бактериальной и животной клетки — являются
чем-то подлинно новым, принципиально отличающимся от исходных клеток, не
содержащих материалы симбионта. Из таких «химер» складывались все более и
более сложные биологические системы, происходило эволюционное обновление.
Доказательством гипотезы
симбиогенеза Л.Маргулис считает присутствие генов в цитоплазме многих
простейших, эти гены являются остаточным генетическим материалом каких-то
вирусов или бактерий, случайно попавших в клетку и «застрявших» в ней.
Обнаружено, что передача внеядерных генов по наследству происходит по другому
механизму, чем передача хромосомных генов. Гены хлоропластов в растительных
клетках и митохондрии в животных клетках также передаются от клетки к ее потомкам
по особым правилам, которые во многом отличаются от правил передачи ядерных
генов.
Следовательно, эукариотные
клетки имеют как минимум две генетические родословные. Миллиарды лет назад эти
важнейшие органеллы были отдельными живыми простейшими организмами. На каком-то
этапе эволюции они вступили в тесный симбиоз с другими простейшими клетками, и
в течение миллионов лет развития образовались эукариоты, владеющие
фотосинтетическим аппаратом; также появились эукариоты, осуществляющие в митохондриях
процесс создания молекул АТФ, позволяющих клетке энергично перемещаться в
поисках пищи.
Образование и эволюция
почвенного покрова также представляют собой пример увеличения информации.
Многообразие почв, дифференцированность их строения, сложность организации этих
биокосных тел есть результат переноса биологической информации на косную
среду. Формирование всех биокосных систем планеты свидетельствует о процессе
повышения сложности их организации, увеличения их разнообразия и,
следовательно, идет с накоплением информации.
Основное выражение
информационного подхода сводится к повышению надежности и устойчивости
биосферы. Устойчивость экосистем и их возможность приспособиться к изменяющимся
абиотическим факторам в значительной степени зависят от сложности экосистем,
т.е. от числа составляющих их видов животных, растений, микроорганизмов, и
взаимодействия между ними и окружающей средой. Эволюция идет в сторону
достижения наибольшей суммы жизни (биомассы, продуктивности), которая возможна
лишь при наибольшем разнообразии видов в экосистемах. Возрастание структурной
сложности биосферы и усложнение взаимодействий между ее компонентами ведут к
повышению ее целостности и уменьшению зависимости от космоса и других
геологических оболочек Земли.
Повышение количества
информации в экосистемах обусловливало не только количественные, но и
качественные преобразования биосферы, было источником дальнейшего ее развития
в связи с усложнением биогеоценотических связей.
Рост информации не был
непрерывным и монотонным процессом. Были периоды резкого сокращения
биологической, а возможно, и неорганической информации. Часть информации постоянно
переходила в ископаемое состояние в виде однородных горных пород. Все
разнообразие месторождений горючих ископаемых не может в какой-либо степени
отразить многообразие организмов, из которых они образовались.
Информационный критерий
позволяет выразить такие тенденции в эволюции биосферы, как возрастание дифференциации
живого, биокосного и косного веществ. Однако он не отражает расширения зоны
распространения жизни, перестройки энергетики биосферы, преобразования
биогеохимических функций, усиления роли живого вещества в выветривании,
почвообразовании, осадкообразовании, в преобразовании атмосферы и гидросферы.
Интегральная характеристика всех этих процессов может быть выработана на основе
изучения исторических преобразований биологического круговорота — главного
механизма биосферы, обеспечивающего его функционирование.
4.Эволюция
биологического круговорота
Увеличение биомассы живого
вещества, возрастание энерговооруженности биосферы и ее информационной
«емкости» являются сторонами единого процесса — эволюции биологического
круговорота. Впервые эту мысль высказал В.И.Вернадский: живое вещество,
избирательно извлекая из окружающей среды элементы и их изотопы, создает и
сохраняет в охваченной ими области общепланетарный биологический круговорот.
Идея получила поддержку в работах почвоведов, географов, геохимиков.
По мнению В. Р. Вильямса (1950), единственный способ придать количественно
ограниченному свойство бесконечного — это придать ему циклическое движение,
заставить его вращаться в круговороте. Этот способ использовала жизнь,
выработав его в процессах длительного отбора и дошлифовки биотических отношений.
Уже развитие примитивных флоро-фаунистических комплексов и начальная их
дифференциация вели к возникновению особых биогенных циклов в сложных
круговоротах вещества и превращениях энергии. Формирующиеся трофические
отношения в зарождающейся биосфере способствовали изыманию элементом из
большого геологического круговорота и вовлечению их в новый, меньший по
сравнению с исходным биологический круговорот. Эти элементы стали вращаться в
круговороте «расширяющейся спиралью». Сложившаяся в биосфере система отношений
продуцентов, консументов и редуцентов обеспечивает удержание зольных элементов
в биологическом круговороте. Постепенно биогенные циклы вещества и энергии
приобрели общепланетарный характер, используя при этом особенности
геологического цикла.
И.П.Герасимов
(1951) показал, что в процессе приспособления растений к условиям низкого
естественного плодородия почв складывался сплошной биоценотический покров,
появлялись все более сложные по структуре и видовому составу биогеоценозы.
Усложнялась пищевая цепь и повышалась интенсивность биологического
круговорота. В его орбиту стали вовлекаться все большие массы вещества, все
более разнообразные по своему составу химические соединения, которые на более
ранних этапах геологической истории свободно мигрировали, не захватываясь
организмами, в поверхностных и грунтовых водах и достигали океана. Возрастающее
влияние живого на среду, расширение сферы существования жизни и вызванные этим
изменения геохимических отношений между океаном и сушей в свою очередь
оказывали воздействие на дальнейшую эволюцию живого. Тем самым усложнение и
совершенствование биологического круговорота представляют собой не только
важнейший результат эволюции биосферы, но и источник ее дальнейшего развития.
![]() Эволюцию биологического круговорота, изменение его отношений с
большим геологическим круговоротом веществ можно рассматривать как общее
усложнение процессов обмена веществ между геологическими оболочками планеты.
Круговорот веществ на начальных стадиях развития Земли рассматривал А.П.Виноградов (1959). Он показал, что
развитие поверхностной оболочки Земли в сторону наиболее устойчивого
динамического равновесия приводит к обмену веществ между отдельными геосферами
и внутри них. Вначале существенную роль играли процессы выноса веществ из недр
Земли в ходе дегазации и выплавления. Особенно интенсивно они шли в архее,
когда происходили крупные колебательные движения земной коры, обширный
орогенез, вулканизм, гранитизация. Громадные массы магматического вещества и
вулканических газов поступали из глубинных слоев на поверхность. Затем зона
обмена веществом между глубинными и поверхностными оболочками резко
сократилась, так как области геосинклиналей сузились. Большое значение приобрел
большой круговорот воды, благодаря которому происходила миграция элементов и
эволюция химического состава геосфер.
Эволюцию биологического круговорота, изменение его отношений с
большим геологическим круговоротом веществ можно рассматривать как общее
усложнение процессов обмена веществ между геологическими оболочками планеты.
Круговорот веществ на начальных стадиях развития Земли рассматривал А.П.Виноградов (1959). Он показал, что
развитие поверхностной оболочки Земли в сторону наиболее устойчивого
динамического равновесия приводит к обмену веществ между отдельными геосферами
и внутри них. Вначале существенную роль играли процессы выноса веществ из недр
Земли в ходе дегазации и выплавления. Особенно интенсивно они шли в архее,
когда происходили крупные колебательные движения земной коры, обширный
орогенез, вулканизм, гранитизация. Громадные массы магматического вещества и
вулканических газов поступали из глубинных слоев на поверхность. Затем зона
обмена веществом между глубинными и поверхностными оболочками резко
сократилась, так как области геосинклиналей сузились. Большое значение приобрел
большой круговорот воды, благодаря которому происходила миграция элементов и
эволюция химического состава геосфер.
Изучение фанерозойных
палеобиосфер и современных геохимических ландшафтов стало основой в понимании
эволюции собственно биологического круговорота (труды Б.Б.Полынова,
Н.И.Базилевич, М.А.Глазовской, В.В.Добровольского, А.И.Перельмана, В. А. Ковды
и др.).
В ходе завоевания жизнью
гидросферы, нижней части тропосферы, верхней части литосферы возрастало
воздействие жизни на миграцию химических элементов. По А. А. Саукову, три главных события повлияли
на эволюцию биологического круговорота: появление фотосинтезирующих растений,
выход растений на сушу, появление человеческого общества. Так, возникновение
наземной флоры вело к накоплению органического вещества в почвах, залежах
торфа, угля и т.д. Формировались некие «органические барьеры», препятствующие
элементам мигрировать с суши в водоемы.
А.И.Перельман
(1975) выделил три этапа в эволюции геохимического ландшафта: абиогенный,
биогенный и культурный. Биогенный ученый разделил на этапы: докембрий, ранний и
поздний палеозой, мезозой и кайнозой. Становление биологического круговорота
он относил к периоду докембрийских геохимических ландшафтов, когда жизнь
пробовала различные варианты биологического круговорота, вовлекая в интенсивную
миграцию разные элементы, пока не возник наиболее целесообразный круговорот.
Со временем нарастала дифференциация биологического круговорота, связанная с
появлением новых циклов элементов, изменением их форм миграции, ускорением
темпа круговорота, совершенствованием способности к максимальному использованию
минерального вещества и воды, к повышению продуктивности ландшафтов. Несмотря
на направленность эволюции биологического круговорота, обусловленную прогрессом
живых организмов, в ней наблюдается и некоторая периодичность. В периоды
тектонического покоя при участии живого вещества в почву поступало больше
угольной и органической кислот, почвенные воды становились более кислыми.
Нарастало выщелачивание и обеднение ландшафтов элементами минерального питания
растений. В эпохи интенсивного орогенеза магматические процессы выносили в
биосферу породы, обогащенные фосфором, калием, кальцием, натрием и другими
макро- и микроэлементами, радиоактивными элементами. Изменение химизма пород и
почв сопровождалось определенными изменениями в биологическом круговороте
веществ.
Высокая зависимость
биологического круговорота от эволюции растительного покрова показана В. А. Ковдой (1985). Возрастание
количества видов с различными биогеохимическими и экологическими функциями
означало усиление биологического круговорота, вовлечение в него все большего
числа химических элементов, а также возрастание роли живого вещества в
процессах выветривания, осадкообразования, преобразования атмосферы,
гидросферы, литосферы. Ковда оценивает появление почвообразовательного
процесса как важнейшее событие в эволюции биосферы, подготовленное появлением
на суше микроорганизмов и растений. Постепенно почвенный покров превратился в
важнейший компонент биосферы, представляя область концентрации процессов
разложения останков животных и растений, а следовательно, и освобождения
громадного количества энергии и накопления питательных веществ. Почва стала
важнейшим аккумулятором вещества и энергии, используемых в биологическом
круговороте.
По В.А. Ковде (1985), появление водорослей повысило роль
биологического круговорота минеральных элементов, так как зольность водорослей
в 3 — 4 раза выше, чем бактерий. Особенно возросла доля натрия, серы и хлора,
наметилась тенденция к увеличению доли кремния и фосфора, получившая дальнейшее
развитие у наземных растений. Размеры биологического круговорота непрерывно
увеличивались как вследствие увеличения количества видов и массы живого, так и
ввиду общего возрастания зольности от хвойных к лиственным древесным породам и
от последних к травянистым. Ковда указывал также на противоречивость эволюции
биологического круговорота: увеличение в тканях растений содержания фосфора,
серы, железа, магния, кальция и особенно калия происходило с колебаниями и
отклонениями. Содержание кремния, достигнув максимума в хвощах, хвойных и
злаках, в дальнейшем уменьшается. Доля хлора в наземных растениях в целом
постепенно снижалась.
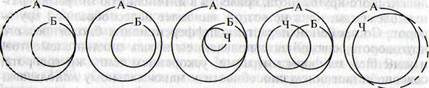
Рис. Стадии развития биосферы
(по М. М. Камшилову, 1974, с изменениями): А — большой абиотический
круговорот; Б — биосфера; Ч — человечество
Биолог М.М. Камшилов (1974), синтезируя дарвинизм с идеями
В.И.Вернадского, говорит о расширении области жизни и усилении давления живых
организмов на среду. Распространение жизни на планете создало предпосылки для
сознательного регулирования обмена вещества и энергии между органическими и
неорганическими компонентами биосферы. Жизнь изначально существовала в форме
биологического круговорота, основанного на взаимодействии синтетиков и
деструкторов. Возник он на базе абиотического круговорота, в недрах которого
стали формироваться циклы органического вещества в виде синтеза и распада
химических соединений углерода. Камшилов выделил следующие основные этапы
эволюции биологического круговорота:
• круговорот, осуществляемый прокариотными
формами жизни;
• возникновение одноклеточных эукариотов;
• усложнение циклической структуры жизни в
результате надстройки трофических отношений из многоклеточных животных,
растений и грибов;
• превращение биологического круговорота в
общепланетарный процесс в связи с завоеванием жизнью суши;
• усложнение и расширение области действия
биологического круговорота с завоеванием доминирующего положения гомойотермными
животными;
• возникновение человеческого общества,
переход к биологическому круговороту, сознательно регулируемому людьми;
• распространение биологического круговорота
на космическое пространство.
Характерная черта эволюции
биологического круговорота заключается в том, что жизнь, извлекая из среды
необходимые вещества и обогащая ее продуктами жизнедеятельности и свободной
энергией, неизбежно изменяет свои условия существования. Она должна постоянно
приспосабливаться к новым, созданным ею же условиям существования. Вместе с ней
неизбежно должен меняться биологический круговорот.
Новые формы жизни возникают
на базе старых и нередко могут существовать лишь вместе с ними. Это не только
усложняет биологический круговорот, но и повышает надежность его функционирования.
Постоянно возрастает темп эволюции в силу усложнения биотических
взаимоотношений, выступающих в роли факторов адаптивных преобразований.
В работах биологов не раз
отмечалось, что эволюция биологического круговорота должна анализироваться с
позиции пищевых цепей. По мнению А. М.
Гилярова (1973), усложнение трофоценотической структуры экосистем не может
идти бесконечно, скорость эволюционных преобразований неизбежно замедляется по
достижении максимально возможного количества трофических уровней. А.М.Уголев (1985) отмечает, что
трофические взаимодействия — единая целостная система, биологический круговорот,
прежде всего трофический процесс. Формирование его началось с редуцентов,
использующих абиогенные органические вещества. По мере истощения его запасов
возникали фотосинтетики и хемосинтетики. Эволюция каждого вида определяется его
приспособленностью к трофической структуре биосферы. Играет роль не только его
обеспеченность источниками питания, но и «поедаемость» каждого члена
трофической цепи, т.е. доступность для других быть источником пищи. Эволюция
трофической системы биосферы направлена на обеспечение высокой эффективности,
экономичности, регулируемости и надежности функционирования биосферы.
Само развитие биосферы
связано с постоянным разрешением противоречия между безграничной способностью
организмов к умножению и ограниченностью материальных ресурсов. Разрешение
этого противоречия способно только на пути создания все более прогрессивных
форм, способных к овладению новыми источниками вещества и энергии и к
рациональному их использованию. Все это усложняло структуру биологического
круговорота.
Ближайшей задачей
эволюционной теории является изучение воздействий микро- и макроэволюции на
исторические преобразования биосферы, на ее биогеохимическую и энергетическую
эволюцию.