Раздел II. ОСНОВНЫЕ ЭТАПЫ ЖИЗНЕННОГО ЦИКЛА РЫБ
Глава 9. Размножение рыб
Размножение – это присущее всем живым организмам свойство воспроизведения себе подобных, обеспечивающее непрерывность и преемственность жизни. Отличается рядом специфических особенностей, обусловленных водным образом жизни. В отличие от наземных животных, зрелые половые клетки рыб выводятся в воду, где происходит оплодотворение икры и дальнейшее ее развитие. Осеменение, оплодотворение и инкубация икры в воде, вне материнского организма, влекут за собой большую гибель потомства на ранних стадиях развития.
Жизненный цикл рыб (рис. 65) от момента оплодотворения до естественной смерти включает ряд периодов.
1. Эмбриональный (зародышевый) – от момента оплодотворения яйца до перехода особи на внешнее питание; эмбрион питается за счет желтка, полученного от материнского организма.
Включает два подпериода: собственно эмбриональный – развитие внутри яйцевой оболочки, и предличиночный (свободного эмбриона) – развитие вне оболочки.
2. Личиночный – характеризуется переходом на внешнее питание с сохранением личиночных органов и признаков.
3. Мальковый – внешнее строение организма приобретает сходство со взрослым. Закладывается чешуя, начинается дифференциация пола, но половые органы неразвиты.
4. Ювенильный (юношеский) – характеризуется усиленным развитием половых желез, но рыбы еще неполовозрелые. Начинают развиваться вторичные половые признаки (если они имеются).
5. Период взрослого организма – с момента наступления половой зрелости. Рыбы обладают всеми признаками, характерными для полностью сформировавшегося организма.
6. Старческий – характеризуется замедлением роста или его прекращением. Рыба теряет способность размножаться.
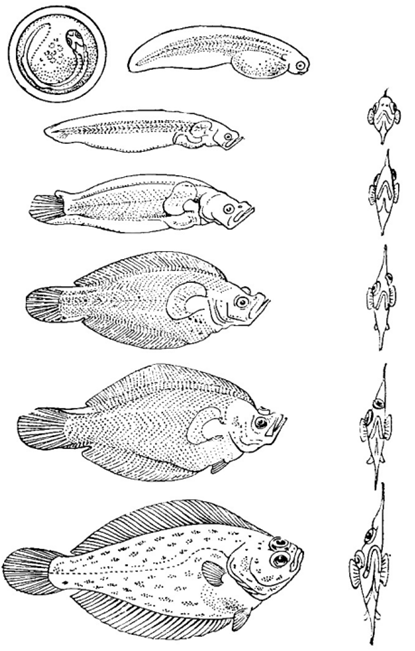
Рис. 65. Схема жизненного цикла камбал
Рыбы размножаются половым путем. В ряде случаев у них существует полиспермия – это проникновение в яйцо при оплодотворении нескольких сперматозоидов. Полиспермия в норме свойственна некоторым группам рыб с внутренним осеменением (например, акулы).
В редких случаях у рыб встречаются:
1) партеногенез (развитие икры без оплодотворения) – развитие икры доходит только до стадии дробления и лишь в исключительных случаях до личинки, доживающей до рассасывания желточного мешка (например, налим, салака).
В большинстве случаев такое развитие не приводит к получению жизнеспособной молоди. У лососей неоплодотворенные икринки, оказавшись в нерестовом бугре вместе с оплодотворенными, нередко развиваются партеногенетически;
2) гиногенез (рождение самок) – сперматозоиды близких видов рыб проникают в яйцо и стимулируют его развитие, однако оплодотворения при этом не происходит. В результате такого размножения в потомстве наблюдаются одни самки, например, встречаются популяции серебряного карася, состоящие из одних самок.
Рыбы, как правило, однополы, но среди них встречаются и гермафродиты. Среди костистых рыб, например, к гермафродитам относятся каменный окунь Serranus scriba, у которого в гонадах развиваются икра и сперматозоиды, но созревание их обычно происходит поочередно, и красный пагель Pagellus erythrinus, у которого в течение жизни происходит изменение (реверсия) пола: у молодых особей гонады функционируют как яичники, у более старших – как семенники.
Оплодотворение у рыб бывает наружное (у большинства рыб) и внутреннее (у хрящевых рыб, у некоторых костистых, например, морской окунь, бельдюга).
В зависимости от вида размножения рыб делят на:
1) яйцекладущих, откладывающих яйца во внешнюю среду (большинство видов);
2) яйцеживородящих, рождающих мальков. У них оплодотворенные яйца задерживаются в задних отделах яйцеводов и развиваются там до вымета молоди (например, белая акула); у некоторых видов, например у ската-хвостокола, стенки задних отделов яйцеводов («матки») имеют даже особые выросты, по которым через брызгальца в ротовую полость эмбрионов поступает питательная жидкость;
3) живородящих, у рыб в задних отделах яйцеводов («матке») образуется нечто сходное с плацентой млекопитающих, а эмбрион получает питательные вещества с кровью матери (например, голубая акула).
Приспособительное значение живорождения и яйцеживорождения рыб заключается в том, что при внутриутробном развитии обеспечивается большая выживаемость молоди.
Развитие женских половых клеток (овогенез) и мужских половых клеток (сперматогенез) у рыб – длительный и сложный процесс. Каждая половая клетка, прежде чем она окончательно созреет, должна пройти в своем развитии ряд стадий. При этом различают два процесса:
1) период до достижения половой зрелости, начиная от возникновения первичных половых клеток и кончая образованием зрелых половых продуктов;
2) периодическое созревание определенной части половых продуктов в течение межнерестового периода (после достижения половой зрелости).
Первый процесс более длительный, чем второй, и у разных видов рыб занимает разное время.
В зависимости от характера размножения рыб делят на:
1) моноцикличных – рыбы после однократного икрометания погибают (речной угорь, тихоокеанские лососи, миноги);
2) полицикличных – рыбы размножаются в течение жизни по нескольку раз (большинство рыб).
Возраст наступления половой зрелости у рыб значительно колеблется – от 1–2 месяцев (например, гамбузия) до 15–30 лет (например, осетровые). Раньше созревают рыбы с коротким жизненным циклом (например, некоторые бычки – в возрасте 1 года), рыбы с продолжительным жизненным циклом становятся половозрелыми значительно позже (например, атлантическая треска – в 7–10 лет, морской окунь – в 12–15 лет).
Возраст полового созревания рыб зависит от видовой принадлежности, условий обитания и в первую очередь от условий откорма.
Наступление половой зрелости у рыб происходит при достижении определенной длины. Как правило, чем лучше питается рыба, тем быстрее она растет, а следовательно, быстрее созревает. Самцы обычно созревают раньше самок. На скорость созревания оказывают влияние и климатические условия. Так, у леща половая зрелость наступает в Аральском море в 3–4 года, на Средней Волге – в 6–7 лет, а в Ладожском озере – в 8–9 лет.
Возраст наступления половой зрелости имеет важное значение для определения размера вылова рыбы и оценки сырьевых ресурсов. У короткоцикловых рыб, созревающих на 2–3 году жизни (например, кильки, сардины), допустимое промысловое изъятие из популяции может составить от 40 до 60 %, у долгоживущих рыб это изъятие должно быть значительно меньше (от 5 до 20 %).
У большинства рыб половой диморфизм (вторичные половые признаки) не проявляется, самки и самцы внешне трудноразличимы. У некоторых видов вторичные половые признаки ярко выражены: самки крупнее самцов, самцам свойственны более яркая окраска, удлиненные плавники и др. Самцы полярной камбалы имеют ктеноидную чешую, самки – циклоидную.
У некоторых рыб в преднерестовый период под влиянием половых гормонов появляется брачный наряд, который исчезает после нереста, например у хариусов краснеют плавники.
У лососей во время нереста серебристая окраска темнеет, на теле появляются черные, малиновые пятна, наблюдаются значительные морфологические изменения (челюсти удлиняются и загибаются, наблюдаются изменения черепного скелета, у горбуши вырастает горб).
Соотношение полов является приспособительным свойством рыб и направлено на обеспечение успешного воспроизводства. У большинства рыб оно близко 1 к 1. Часто самки преобладают у рыб с низкой плодовитостью, у которых самцы охраняют икру нескольких самок, например у некоторых бычков.
Соотношение полов у рыб может меняться под воздействием различных факторов. Например, воздействие высокой температуры на мальков зеленого меченосца приводит к преобладанию самцов.
Гормональные препараты также могут способствовать изменению нормальных соотношений полов у рыб. В рыбоводстве при разведении товарной рыбы может иметь место направленное изменение пола путем добавки в корм стероидных гормонов.
Среди рыб различают:
1) моногамов – с одной самкой обычно нерестится один самец (например, сёмга);
2) полигамов – на одну самку приходится три и более самцов (например, сазан) или один самец обеспечивает оплодотворение икры нескольких самок (например, колюшка).
У большинства рыб самцы первыми подходят к нерестилищам. Затем на местах нереста появляются самки. Соотношение самок и самцов на нерестилищах в среднем обычно 1:1.
Во время нереста самки выметывают икру, а самцы осеменяют ее. Соприкосновение спермы с икрой называется осеменением.
Самки после вымета икры уходят с нерестилищ. Исключение составляют лишь самки некоторых видов рыб. У большинства рыб самцы, после осеменения икры первых подошедших к местам нереста самок, остаются на нерестилищах. Они принимают участие в нересте с другими самками, которые подходят на нерестилища позже. В течение нерестового периода один самец может осеменять икру нескольких самок.
Продолжительность нереста одного вида рыб зависит от типа икрометания (рыбы с порционным нерестом имеют более длительный нерестовый период, чем рыбы с единовременным нерестом), от структуры нерестового стада (например, при многовозрастной структуре стада нерест растянут во времени), от наличия в популяции экологических групп (т. е. разные экологические группы могут иметь разное время нереста), от температуры воды (например, понижение температуры воды в период нереста приостанавливает этот процесс и тем самым увеличивает его продолжительность).
В зависимости от сроков размножения различают рыб:
– весенненерестующих (например, щука, окунь);
– летненерестующих (например, сазан, осётр);
– осенне-зимненерестующих (например, сёмга, навага).
Сроки размножения каждого вида, а также сроки выклева личинок и развития молоди связаны с обеспеченностью их пищей.
По продолжительности периода икрометания выделяют две группы рыб:
– с единовременным нерестом – вся икра созревает одновременно (например, щука, сиг);
– с порционным икрометанием – икра созревает и выметывается порциями в течение длительного времени (например, хамса, уклейка, каспийские сельди, сом, из морских – минтай).
У одного и того же вида в одном водоеме может быть единовременное, а в другом – порционное икрометание. Так, у леща в Аральском море отмечено порционное икрометание, а в северных водоемах (например, Онежское озеро) он выметывает икру единовременно.
Порционное икрометание является приспособлением вида к воздействию факторов среды и способствует увеличению плодовитости, большей выживаемости икры и личинок, лучшему питанию молоди, благодаря равномерному использованию кормовой базы.
Для оценки степени зрелости половых продуктов у рыб применяются шкалы зрелости, из которых наиболее распространенной для полицикличных рыб с единовременным икрометанием является шестибалльная шкала:
I стадия – ювенильная (juveniles), неполовозрелые рыбы. Половые клетки яичников невозможно различить невооруженным глазом, и пол визуально не определяется. Яичники и семенники имеют вид тонких прозрачных тяжей желтоватого или розоватого цвета.
II стадия – созревающие особи. Икринки очень мелкие и видны только под лупой. Яичники прозрачны и бесцветны, вдоль них проходит крупный кровеносный сосуд. Семенники увеличиваются в размерах, теряют прозрачность и имеют вид округлых тяжей сероватого или бледно-розового цвета.
III стадия – особи, у которых половые железы далеки от зрелости, но сравнительно хорошо развиты. Яичники заполняют от 1/3 до 1/2 всей брюшной полости, наполнены непрозрачными икринками, ясно различимыми невооруженным глазом. Семенники плотные, упругие. При надавливании из семенников нельзя выделить жидких молок. Цвет их от розовато-серого до желтовато-белого.
IV стадия – особи, у которых половые железы почти достигли полного развития. Яичники и семенники имеют наибольшую величину и заполняют до 2/3 всей брюшной полости. Икринки округлые, прозрачные и при надавливании вытекают. Семенники мягкие, белого цвета, наполнены жидкими молоками, при надавливании вытекают.
V стадия – текущие особи. Икра и молоки настолько зрелые, что свободно вытекают при легком надавливании на брюшко.
VI стадия – отнерестившиеся особи (выбой). Половые продукты выметаны полностью. Гонады в виде спавшихся мешков. В яичниках могут наблюдаться оставшиеся икринки, в семенниках – остатки спермы. Яичники и семенники воспалены, темно-красного цвета. Через некоторое время после размножения яичники и семенники переходят во II стадию зрелости.
У рыб с порционным икрометанием (рис. 66) стадия зрелости определяется состоянием той порции, которая наиболее развита и раньше всех будет выметана. После вымета первой порции яичники переходят не в VI стадию, как у рыб с единовременным икрометанием, а в IV или III, и эти стадии зрелости обозначаются VI–IV или VI–III. После завершения всего нерестового периода, состояние яичника оценивается как находящееся в VI, а затем во II стадии. Если же оставшиеся ооциты (резерв будущего года) вступают в рост уже на VI стадии, то яичник из VI стадии переходит в III стадию и обозначается VI–III.
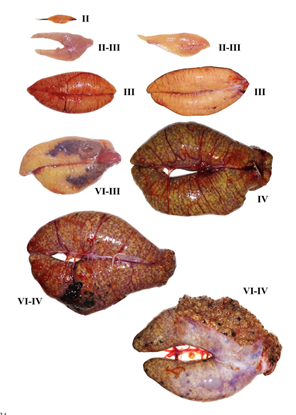
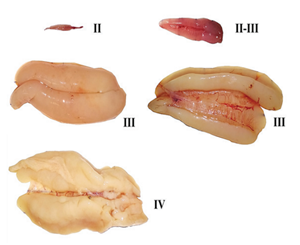
Рис. 66. Яичники (слева) и семенники (справа)
северного однопёрого
терпуга на разных стадиях зрелости
При оценке степени зрелости гонад рыб используют коэффициент и индекс зрелости.
Коэффициент зрелости – это отношение массы гонад к массе тела рыбы (в %). У рыб с весенне-летним нерестом коэффициент зрелости наиболее высок весной, уменьшается летом, начинает снова увеличиваться осенью (например, сазан, судак, вобла). У рыб с осенне-зимним нерестом наиболее высокий коэффициент зрелости осенью (например, сиг).
Индекс зрелости – это процентное отношение коэффициента зрелости гонад, вычисленное в отдельные периоды созревания гонад, к максимальному коэффициенту зрелости.
Строение половых продуктов. Икринки рыб различаются формой, размером, цветом, наличием жировых капель, строением оболочки. У рыб икринки обычно имеют шаровидную форму, но встречаются и другие. Например, у представителей сарганообразных – шаровидная икринка с нитевидными выростами; у бычковидных – грушевидные икринки на нижнем конце снабжены розеткой нитей для прикрепления к субстрату; у анчоусовых – эллипсовидные икринки. Величина икринок, как и другие морфологические признаки, является стабильным признаком вида. Крупные рыбы откладывают икру большего диаметра. Размеры икринок зависят от содержания в них питательного вещества (то есть желтка) и значительно колеблются.
Среди костистых рыб наиболее мелкие икринки встречаются у камбалы-лиманды, самые крупные – у лососевых (например, кета). Хрящевые рыбы имеют самые крупные яйца. Так, у разнозубых акул длиной 1,5 м длина яйцевой капсулы около 10 см. Развитие эмбрионов у некоторых из них длится очень долго – от 18 до 22 месяцев (например, акула катран).
Окраска икринок специфична для каждого вида. Икра, развивающаяся в менее благоприятных кислородных условиях, обычно окрашена интенсивнее. У ряпушки икра желтая, у лососей – оранжевая, у щуки – темно-серая, у сазана – зеленоватая, у терпугов – изумрудно-зеленая, голубая, розовая и фиолетовая. Желтоватый и красноватый цвет икры объясняется наличием дыхательных пигментов (каротиноидов). Пелагические икринки, развивающиеся при достаточном содержании кислорода, пигментированы слабо.
Икринки многих рыб содержат одну или несколько жировых капель, которые обеспечивают их плавучесть.
Снаружи икринки покрыты оболочками:
1) первичная – желточная (лучистая) оболочка, образована самим яйцом, пронизана многочисленными порами, по которым в яйцо поступают питательные вещества во время его развития в яичнике. У некоторых видов эта оболочка двухслойная (например, осетровые);
2) вторичная – студенистая, липкая (развивается над первичной оболочкой), с разнообразными выростами для прикрепления яиц к субстрату.
На анимальном полюсе обеих оболочек расположен особый канал – микропиле, по которому при оплодотворении сперматозоид проникает в яйцо. У костистых имеется один канал, у осетровых их может быть несколько;
3) третичная – роговая (у хрящевых рыб и миксин) и белковая (только у хрящевых).
У миног, как и у костистых рыб, икринки мелкие, у миксин они эллипсовидной формы длиной 2–3 см (рис. 67). На роговой оболочке миксин имеются крючкообразные отростки, с помощью которых яйца прикрепляются друг к другу и к подводным предметам. Роговая оболочка хрящевых рыб значительно больше самого яйца. Часто от нее отходят роговые нити, с помощью которых яйцо прикрепляется к субстрату.
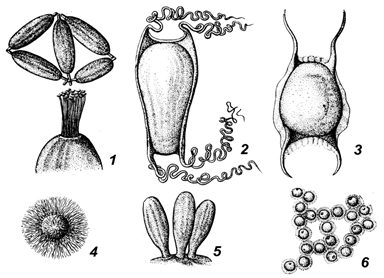
Рис. 67. Форма яйцеклеток у различных видов рыб: 1 – миксина; 2 – акула; 3 – скат; 4 – сарган; 5 – бычок; 6 – окунь
В сперматозоиде различают головку, среднюю часть и хвост (рис. 68). Форма головки может быть шаровидной (у большинства костистых рыб), палочковидной (у осетровых и некоторых костистых), копьевидной (у двоякодышащих), цилиндрической (у акуловых, кистеперых). В головке помещается ядро. Впереди ядра у акулообразных, осетровых и некоторых других рыб располагается акросома – органоид сперматозоида, расположенный в передней части его головки. Обычно он чашевидный или копьевидный. У костистых ее нет.
![]()
![]()

Рис. 68. Сперматозоиды
костистых (А) и хрящевых (Б) рыб: 1 – карась;
2 – щука; 3 – подкаменщик (вид головки с уплощенной стороны и
сбоку);
а – акросома; б – головка; в – хвост
Сперма, выделяемая самцом, состоит из сперматозоидов, погруженных в спермиальную жидкость, сходную по составу с физиологическим раствором. В спермиальной жидкости сперматозоиды неподвижны. При соприкосновении с водой их активность резко возрастает. Встретив икринки, они проникают в них через микропиле, после чего происходит оплодотворение.
Продолжительность активности сперматозоидов зависит от солености и температуры воды. В соленой воде они живут значительно дольше – до нескольких суток (тихоокеанские сельди), в пресной – не более 1–3 минут (у большинства рыб – карповые, лососевые, окуневые).
У одного и того же самца сперматозоиды качественно неодинаковые и различаются по величине. При центрифугировании выделяют: мелкие (легкие), средние (промежуточные) и крупные (тяжелые) сперматозоиды.
Среди крупных сперматозоидов в большом количестве встречаются X-гаметы, среди мелких – Y-гаметы. Таким образом, из икры, оплодотворенной крупными сперматозоидами, рождаются преимущественно самки, мелкими – самцы. Это имеет важное значение при искусственном разведении ценных видов рыб.
Начало нереста рыб зависит от различных факторов – готовность половых продуктов, температура и соленость воды, наличие нерестового субстрата.
Из абиотических факторов важным является температура воды. Каждому виду в период размножения свойственны различные оптимальные температуры. При отрицательной – размножаются сайка, наваги, арктические и антарктические рыбы. Минимальная температура воды, при которой возможен нерест, для наваги ‒ 2,3 °С, трески – 3,6 °С, атлантической сельди – 4,5 °С, сазана – 13 °С. У многих карповых рыб наиболее интенсивный нерест наблюдается при температуре 18–20 °С и выше.
Рыбы откладывают икру в разных условиях – одни виды нерестятся в приливно-отливной зоне (например, мойва), другие – в океанической пелагиали на глубинах более 1 тыс. м (например, угорь). Подавляющее большинство морских рыб нерестится в относительно прогреваемых районах прибрежной зоны на глубинах менее 500 м, там, где высокая концентрация кормовых организмов и личинки будут обеспечены пищей.
В нерестовый период у самок рыб, у которых половые железы находятся в IV стадии зрелости, под воздействием гонадотропного гормона гипофиза завершается оогенез и происходит овуляция (икринки выходят из фолликул). Овуляция (выброс яйцеклетки из яичника навстречу сперматозоиду) происходящий в среднем на 14 день цикла, у рыб осуществляется только при определенных условиях внешней среды, среди которых важное значение имеют температура воды, гидрохимический режим, наличие или отсутствие течения воды, присутствие самцов, а для многих рыб необходим нерестовый субстрат. Если после наступления для данного вида рыб нерестовых температур хотя бы один из перечисленных факторов среды неудовлетворителен, то овуляция не происходит, ооциты подвергаются резорбции, а фолликулы – атрезии (зарастанию). Для рыб это явление имеет большое приспособительное значение, так как их яичники освобождаются от ооцитов старшей генерации и начинают новый цикл развития.
После овуляции происходит брачный акт, который называется нерестом, а то место, где он совершается, – нерестилищем.
Выделяют икру: пелагическую (плавающую) и донную (демерсальную), которая откладывается на грунт и донную растительность.
В зависимости от места нереста у рыб встречаются следующие экологические группы:
1) литофилы (рис. 69) – откладывают икру на каменисто-галечный грунт (например, осетровые, лососевые, кутум, шемая, голавль, подуст);

Рис. 69. Кладка икры тихоокеанских лососей
2) фитофилы (рис. 70) – откладывают икру на растения и водоросли (например, вобла, лещ, сазан, карась, окунь, тихоокеанская сельдь);
3) псаммофилы – откладывают икру на песок (например, пескарь);
4) пелагофилы (рис. 71) – откладывают плавающую икру в толщу воды (например, кильки, хамса, атлантическая треска, чехонь, белый амур, толстолобик);

Рис. 70. Кладки икры фитофильных рыб: скалярия (слева) и окунь (справа)
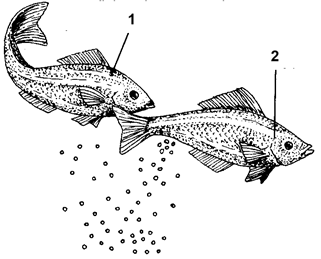
Рис. 71. Схема нереста пелагофильных рыб: 1 –
самец, ожидающий
окончания икрометания, 2 – самка, выметывающая икру в толщу воды
5) остракофилы и карцинофилы (рис. 72) – откладывают икру в раковины двустворчатых моллюсков (например, горчаки) или под панцирь крабов (некоторые морские слизни). Когда подходит время метать икру, у самки появляется длинный яйцеклад, помогающий ввести икринки между створками раковины в жабры моллюска или под панцирь краба.
Большинство рыб не заботятся о своем потомстве. Однако ряд видов сооружают различные гнезда, охраняют икру и личинок. Тихоокеанские и атлантические лососи хвостом вырывают в грунте гнезда длиной до 2–3 м, шириной 1,5–2 м, откладывают в них икру, оплодотворяют ее и засыпают гравием. Самец колюшки сооружает гнездо из растительных остатков в виде муфточки и охраняет икру (рис. 73А). Самец судака расчищает на дне место для будущей кладки икры, затем охраняет ее, очищает от ила, смывая его сильными движениями грудных плавников. Если кладка остается без сторожевого самца, то охрану продолжает другой. Лабиринтовые рыбы строят гнездо из пузырьков воздуха, обволакивая их клейкими выделениями изо рта. Самец пинагора охраняет кладку икры (рис. 73Б), отложенную в литоральной зоне, и при обсыхании поливает икру изо рта водой.
![]()
![]()

Рис. 72. Откладка икры: А – карцинофильной
(морской слизень)
и Б – остракофильной (горчак) рыбами

Рис. 73. Примеры заботы о потомстве у рыб: А
– трёхиглая колюшка,
Б – пинагор
У рыб различают абсолютную (индивидуальную), популяционную, относительную и рабочую плодовитость.
Абсолютная (индивидуальная) плодовитость – это количество икры, откладываемое самкой в течение одного нерестового периода.
Популяционная плодовитость – это количество икры, которое может быть отложено популяцией за один нерестовый сезон.
Плодовитость является приспособительным свойством вида и значительно колеблется. Наиболее низкая плодовитость у хрящевых рыб. Так, у акул она колеблется от 2 до 100 мальков. У костистых рыб наименьшую плодовитость имеет европейская бельдюга (от 10 до 400 мальков), наибольшую – рыбы, выметывающие пелагическую икру (например, луна-рыба – до 300 млн икринок).
Количество и качество икры зависят от массы тела, возраста, жирности и факторов среды. С ростом рыбы и увеличением ее массы тела абсолютная плодовитость повышается.
Рыбы способны регулировать плодовитость в зависимости от изменяющихся условий среды. Большая плодовитость вырабатывается у видов в условиях более интенсивной смертности. Изменение абсолютной плодовитости регулируется через изменение обеспеченности пищей.
Улучшение условий откорма приводит к ускорению темпа роста, а следовательно, к более высокой плодовитости одноразмерных рыб. В связи с этим плодовитость одного вида в разных водоемах различна, отражает условия существования рыб и направлена на обеспечение определенной величины пополнения.
Относительная плодовитость – это количество икринок, приходящееся на 1 г массы тела самки.
Рабочая плодовитость представляет собой количество икринок, получаемое от одной самки для рыбоводных целей.
У пеляди, например, она составляет около 70 % абсолютной (индивидуальной) плодовитости.
В ряде случаев рассчитывают видовую абсолютную плодовитость и популяционную плодовитость.
На воспроизводительную способность рыб сильное влияние оказывает их возраст, т. к. качество половых продуктов на протяжении жизни различно. У большинства видов наиболее высококачественное потомство получается от рыб среднего возраста. Молодые и очень старые особи дают менее жизнестойкое потомство.
У рыб длительность инкубационного периода колеблется от нескольких часов (например, данио) до 22 месяцев (например, колючая акула). Для инкубации икры требуется определенное количество тепла, выражаемое в градусо-днях. Эта величина меняется в зависимости от температуры воды: при повышении (в пределах, свойственных данному виду) развитие икры протекает быстрее. У карповых рыб икра развивается в течение 3–6 дней, у наваги – 3–4 месяцев, у лососей – до 5–6 месяцев.
Численность популяции во многом зависит от выживаемости эмбрионов и обеспеченности пищей личинок на этапе перехода на активное питание. На эти периоды приходится наибольшая смертность по сравнению со всеми другими периодами жизни рыбы.
Основными факторами, определяющими выживаемость эмбрионов и предличинок, являются температура воды, соленость, газовый режим, ветер, волнения.
Температура, при которой эмбриогенез протекает наиболее нормально, называется температурным оптимумом. Повышение температуры ускоряет эмбриогенез, а понижение – замедляет этот процесс.
Температура, ниже которой эмбриогенез не происходит, называется температурным порогом. Однако пороговая температура не приводит к гибели зародышей при воздействии в течение ограниченного промежутка времени. При восстановлении оптимальной температуры эмбриогенез возобновляется и протекает нормально.
Температура, выше которой эмбриогенез прекращается и зародыш погибает, называется температурным максимумом.
У некоторых рыб развитие личинок проходит с метаморфозом (рис. 74). Например, у камбал, речного угря, луны-рыбы, миног. Личинки камбалы имеют симметричное строение: они плавают в верхних слоях воды спиной кверху, со временем постепенно опускаются в более глубокие слои воды и утрачивают двустороннюю симметрию; один глаз у них переходит на другую сторону тела, и после завершения метаморфоза молодые камбалы начинают вести придонный образ жизни.
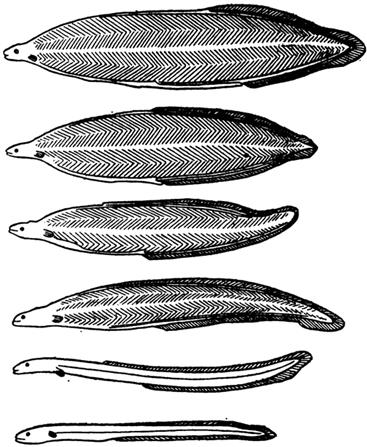
Рис. 74. Схема метаморфоза личинки (лептоцефала) угря в молодую особь
Нерестилища рыб – это места икрометания рыб, благоприятные для развития икры и обитания выклюнувшихся личинок.
Естественными нерестилищами рыб, выметывающих клейкую икру, служат участки водоемов с каменисто-галечным дном или заросшие растительностью. Нерестилища рыб с неклейкой (плавучей) икрой – это участки рек (или морей), обычно с быстрым течением.
Фитофильные рыбы (карп, сазан, лещ, карась, плотва), имеющие клейкую икру, откладывают ее на растительности в стоячих водах, на некотором расстоянии от дна. В этих условиях икра хорошо аэрируется и поэтому исключается опасность гибели ее от недостатка кислорода. Вылупившиеся из икры личинки с помощью клейкого вещества, выделяемого железками, расположенными на голове, подвешиваются к растениям и в таком состоянии, малозаметные для врагов, остаются до конца рассасывания желточного мешка, после чего начинают активную жизнь.
Литофильные рыбы (осетровые, голавль, жерех) тоже имеют клейкую икру, прочно приклеивающуюся к каменисто-галечному субстрату перекатов рек, на которых эти рыбы нерестятся. Икра развивается на дне, где при наличии течения создаются хорошие кислородные условия.
Пелагофильные рыбы (сельди, чехонь, белый амур, толстолобик) выметывают икру непосредственно в толще воды. Плавучесть икринок у таких рыб обеспечивается набуханием оболочек и сильным увеличением объема икринок в воде. В стоячей воде икринки тонут, но даже слабое течение или волнение подхватывают их, и они плывут. Личинки, увлекаемые течением, развиваются также в толще воды.
С целью улучшения условий размножения ценных промысловых рыб, путь к нерестилищам которых перегорожен гидротехническими сооружениями (обычно в верховьях рек), устраивают искусственные нерестилища.
Нерестилища для рыб, откладывающих икру на твердые грунты (например, осетровых, лососей, форелей, сигов), устраивают в руслах рек из гальки или щебня в виде полос или гряд, расположенных обычно перпендикулярно к берегу на разных глубинах (для лососей – также специальные нерестовые каналы).
Для рыб, откладывающих икру на растения (например, карпа, сазана, леща, карася, судака, воблы), делают стационарные или плавучие нерестилища из веток можжевельника, ели, корневищ тростника, небольших деревьев или кустов (затопляемых в прибрежных зонах). Стационарные нерестилища закрепляют на дне на мелких местах, плавучие (деревянные рамы с привязанными к ним пучками веток) устанавливают на относительно глубоких местах с помощью якорей. Сооружение искусственных нерестилищ имеет целью помочь рыбе провести нерест в наиболее благоприятных условиях и тем самым обеспечить нормальное развитие икры и выход молоди. Искусственные нерестилища служат также для сбора и уничтожения икры сорных рыб, т. е. видов рыб, не представляющих хозяйственной ценности, которые могут негативно влиять на популяции ценных пород, конкурируя за корм, поедая икру и мальков, перенося различные заболевания.
Вопросы для самоконтроля
1. Какие периоды включает жизненный цикл рыб?
2. Что такое полиспермия, партеногенез, гиногенез?
3. Чем отличаются яйцекладущие, яйцеживородящие и живородящие рыбы?
4. На какие группы делят рыб в зависимости от сроков размножения?
5. Чему способствует порционное икрометание?
6. Какие стадии входят в шестибалльную шкалу для оценки степени зрелости половых продуктов у рыб с единовременным икрометанием? Дайте их краткое описание.
7. Что такое коэффициент и индекс зрелости?
8. Какие оболочки икринки у рыб? Дайте их краткое описание.
9. Какие части у сперматозоида рыб?
10. Что такое абсолютная (индивидуальная), популяционная, относительная и рабочая плодовитости?