Глава 6. Нервная система и органы чувств рыб
Нервная система у рыб (рис. 42) представлена центральной нервной системой и связанной с ней периферической и вегетативной (симпатической) нервной системой. Нервная система рыб по сравнению с нервной системой высших позвоночных характеризуется рядом примитивных черт.
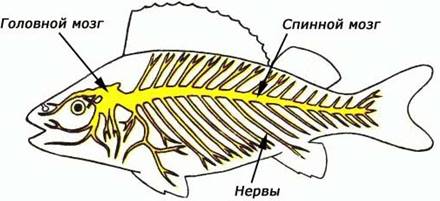
Рис. 42. Схема нервной системы рыб
Центральная нервная система состоит из головного и спинного мозга. Она имеет вид нервной трубки, тянущейся вдоль туловища. Ее часть, лежащая над позвоночником и защищенная верхними дугами позвонков, образует спинной мозг, а расширенная передняя часть, окруженная хрящевым или костным черепом, составляет головной мозг. Трубка имеет внутри полость (невроцель), представленную в головном мозге желудочками мозга. В толще мозга различают серое вещество, слагающееся из тел нервных клеток и коротких отростков (дендритов), и белое вещество, образованное длинными отростками нервных клеток нейритами, или аксонами.
Общая масса мозга у рыб мала: в среднем она составляет у современных хрящевых рыб от 0,06 до 0,44 % от массы тела, у костистых – от 0,02 до 0,94 %, а, например, у птиц – от 0,2 до 8,0 %, у млекопитающих – от 6,3 до 3,0 %.
Отделы головного мозга (рис. 43) у рыб располагаются линейно. В нем выделяют передний мозг, промежуточный, средний, мозжечок (задний) и продолговатый, переходящий в спинной мозг. Полости переднего, промежуточного и продолговатого мозга называются желудочками, полость среднего мозга – сильвиевым водопроводом (она соединяет полости промежуточного и продолговатого мозга, т. е. третий и четвертый желудочки).
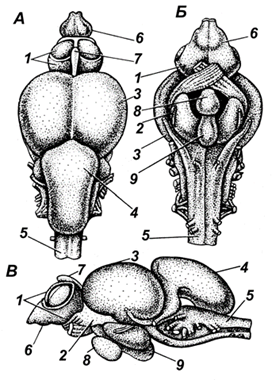
Рис. 43. Строение головного мозга лосося: А – вид сверху; Б
– вид снизу;
В – вид сбоку; 1 – передний мозг; 2 – промежуточный мозг; 3
– средний мозг (зрительные доли); 4 – мозжечок; 5 – продолговатый
мозг;
6 – обонятельные доли; 7 – эпифиз; 8 – гипофиз; 9 –
сосудистый мешок
Передний мозг рыб не образует полушарий. К нему прилегают обонятельные луковицы (первичный обонятельный центр) или непосредственно (у большинства видов), или через обонятельный тракт (у карповых, сомовых, тресковых).
В крыше переднего мозга нет нервных клеток. Серое вещество в виде полосатых тел сосредоточено главным образом в основании и обонятельных долях, выстилает полость желудочков и составляет главную массу переднего мозга. Волокна обонятельного нерва связывают луковицу с клетками обонятельной капсулы.
Передний мозг является центром обработки информации, поступающей от органов обоняния. Благодаря своей связи с промежуточным и средним мозгом он участвует в регуляции движения и поведения. В частности, передний мозг принимает участие в формировании способности к таким актам, как икрометание, охрана икры, образование стаи и т. д.
У костистых рыб передний мозг представлен двумя структурами – мантией (плащ) и гиппокампом (первичная мантия). Мантия хорошо развита у рыб, для которых обонятельная сенсорика выходит на первый план во взаимодействиях с внешней средой, например у европейского угря.
Промежуточный мозг состоит из трех основных частей: дорзальной (эпиталамус, эпифиз), средней (таламус) и вентральной (подбугорье, или гипоталамус).
От таламуса отходят зрительные нервы, образующие хиазму (перекрест, т. е. часть волокон правого нерва переходит в левый нерв и наоборот). Хиазма позволяет создавать в мозге единую картину, что важно для ориентации во время движения, отыскивания и захвата пищи, сохранения стаи и других задач.
На нижней стороне гипоталамуса имеется воронка, к которой прилегает гипофиз. Эпифиз развивается в верхней части промежуточного мозга. Гипофиз и эпифиз являются железами внутренней секреции. Они осуществляют гормональную регуляцию обменных процессов.
Промежуточный мозг выполняет многочисленные функции. Он воспринимает раздражения от сетчатки глаза, участвует в координации движений, в переработке информации от других органов чувств.
Средний мозг – наибольший по объему. Он имеет вид двух полушарий (зрительные доли). Зрительные доли являются первичными зрительными центрами, воспринимающими возбуждение. Из этих долей берут начало волокна зрительного нерва. В среднем мозге обрабатываются сигналы, идущие от органов зрения и равновесия; здесь помещаются центры связи с мозжечком, продолговатым и спинным мозгом. У костистых рыб в среднем мозге формируется примитивная кора (тектум), в ней происходит синтез восходящей информации.
Мозжечок расположен в задней части мозга и может иметь форму или маленького бугорка, прилегающего сзади к среднему мозгу, или большого мешковидно-вытянутого образования, примыкающего сверху к продолговатому мозгу. Особенно большого развития достигает мозжечок у сомов. Мозжечок является центром всех моторных иннервации при плавании, схватывании пищи. Он обеспечивает координацию движений, поддержание равновесия, мышечную деятельность, связан с рецепторами органов боковой линии.
Продолговатый мозг, пятый отдел головного мозга, без резкой границы переходит в спинной мозг. Полость продолговатого мозга – четвертый желудочек продолжается в полость спинного мозга – невроцель. Значительная масса продолговатого мозга состоит из белого вещества.
От продолговатого мозга отходит большая часть (6 из 10) черепно-мозговых нервов. Он является центром регуляции деятельности спинного мозга и вегетативной нервной системы. В нем располагаются наиболее важные жизненные центры, регулирующие деятельность дыхательной, скелетно-мышечной, кровеносной, пищеварительной, выделительной систем, органов слуха и равновесия, вкуса, боковой линии, электрических органов у имеющих их рыб. Поэтому при разрушении продолговатого мозга, например при перерезке туловища позади головы, наступает быстрая смерть рыбы. Через приходящие в продолговатый мозг спинномозговые волокна осуществляется связь продолговатого и спинного мозга.
Передний мозг (и обонятельные доли) относительно сильнее развит у хрящевых рыб (акул и скатов) и слабее – у костистых. У малоподвижных, например донных, рыб мозжечок мал, но сильнее развиты передний и продолговатый отделы мозга в соответствии с большой ролью обоняния и осязания в их жизни (камбалы). У хорошо плавающих рыб (пелагических, питающихся планктоном, или хищников), наоборот, гораздо большее развитие получают средний мозг (зрительные доли) и мозжечок (в связи с необходимостью быстрой координации движения). Рыбы, обитающие в мутной воде, имеют маленькие зрительные доли, небольшой мозжечок. Слабо развиты зрительные доли у глубоководных и слепых рыб.
Спинной мозг является продолжением продолговатого мозга. Он имеет форму округлого тяжа и проходит в канале внутри верхних нервных дуг позвонков по всей длине позвоночника рыбы. В спинном мозге серое вещество расположено внутри, а белое – снаружи. Спинной мозг сегментирован: от него метамерно соответственно каждому позвонку, отходят спинно-мозговые нервы, иннервирующие поверхность тела, туловищные мышцы, а благодаря соединению спинномозговых нервов с ганглиями симпатической нервной системы – и внутренние органы.
Функции спинного мозга – рефлекторная и проводящая. В спинном мозге костистых рыб имеется секреторный орган – урогипофиз, клетки которого вырабатывают гормон, участвующий в водном обмене.
К периферической нервной системе относятся нервы, отходящие от головного и спинного мозга к органам. От головного мозга отходят 10 пар черепно-мозговых нервов:
1) обонятельный нерв от чувствующего эпителия обонятельной капсулы доводит раздражения до обонятельных луковиц переднего мозга;
2) зрительный нерв тянется до сетчатки глаза от зрительных бугров промежуточного мозга;
3) глазодвигательный нерв иннервирует мышцы глаза, отходя от дна среднего мозга; управляет движением глаза – мышцами, которые управляют положением глаза в глазнице.
4) блоковый нерв тянется от крыши среднего мозга к верхней косой мышце глаза;
5) тройничный нерв, отходящий от боковой поверхности продолговатого мозга и дающий три основные ветви: глазничную, верхнечелюстную и нижнечелюстную;
6) отводящий нерв тянется от дна продолговатого мозга к прямой мышце глаза;
7) лицевой нерв отходит от продолговатого мозга и дает многочисленные разветвления к мускулатуре подъязычной дуги, слизистой ротовой полости, коже головы (в т. ч. боковой линии головы);
8) слуховой нерв связывает продолговатый мозг и слуховой аппарат;
9) языкоглоточный нерв идет от продолговатого мозга к глотке, иннервирует слизистую глотки и мускулатуру первой жаберной дуги;
10) блуждающий нерв наиболее длинный, связывает продолговатый мозг с жаберным аппаратом, кишечным трактом, сердцем, плавательным пузырем, боковой линией.
Вегетативная нервная система в основе имеет многочисленные ганглии и нервы, иннервирующие мышцы внутренних органов и кровеносных сосудов сердца. Она действует в определенной степени автономно, независимо от центральной нервной системы и определяет непроизвольную, автоматическую деятельность внутренних органов даже в том случае, если ее связь с центральной нервной системой нарушена.
Вегетативная нервная система у хрящевых рыб представлена разобщенными ганглиями, лежащими вдоль позвоночника. Клетки ганглиев своими отростками контактируют со спинномозговыми нервами и внутренними органами.
У костистых рыб ганглии вегетативной нервной системы соединяются двумя продольными нервными стволами. Соединительные ветви ганглиев связывают вегетативную нервную систему с центральной. Взаимосвязи центральной и вегетативной нервной систем создают возможность некоторой взаимозаменяемости нервных центров.
Реакцию организма рыбы на внешние и внутренние раздражения определяет рефлекс. У рыб можно выработать условный рефлекс на свет, форму, запах, вкус, звук. По сравнению с высшими позвоночными у рыб условные рефлексы образуются медленнее, а угасают быстрее.
Органы чувств – это сенсорные системы рыб, которые позволяют им воспринимать разнообразную информацию об окружающем мире и сигнализировать о внутреннем состоянии организма.
Органы чувств рыб: зрение, слух, обоняние, осязание, вкус, боковая линия, вестибулярная система, электрорецепция, хеморецепция, механорецепция, магниторецепция.
Зрение. Органы зрения информируют живой организм одновременно о месте расположения, контурах, величине, подвижности или неподвижности объекта, направлении движения и его удаленности.
Экспериментально установлено, что видимый для рыб спектр электромагнитного излучения лежит в той же зоне, что и у высших позвоночных. Однако водная среда определенным образом изменяет диапазон восприятия электромагнитного излучения. Так, инфракрасные (ИК) лучи не проникают в воду, поэтому не воспринимаются глазом рыб. Ультрафиолетовые (УФ) лучи также рыбами не воспринимаются.
В разных водоемах при различной их освещенности (характер облачности, расположение солнца относительно горизонта, сезон года) оптическое поле одной и той же особи будет иметь разные характеристики.
Зрительная система рыб включает парные глаза, расположенные на голове в большинстве случаев симметрично по ее бокам. Исключение составляют некоторые морские виды, например скаты или камбалы, у которых органы зрения расположены на одной, «верхней» стороне. изменять диаметр только в небольших пределах.
Век у рыб, как правило, нет. Лишь акулы имеют мигательную перепонку, закрывающую глаз, как занавеска, и некоторые сельди и кефали имеют жировое веко (прозрачную пленку, закрывающую часть глаза). У рыб отсутствуют слезные железы, так как, в отличие от наземных животных, им нет необходимости смачивать глазное яблоко.
Строение глаза рыб. Глаза рыб (рис. 44) устроены примерно, как у человека: свет попадает на радужку, проходит через зрачок и преломляется на хрусталике. Хрусталик подает свет на сетчатку, состоящую из фоторецепторов двух видов (палочек и колбочек). На сетчатке возникает изображение, которое видит рыба.
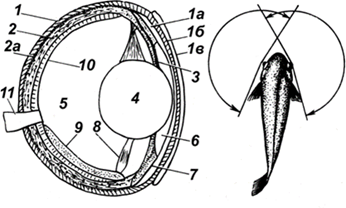
Рис. 44. Строение глаза и поле зрения рыб: 1 – склера; 1а,
1б, 1в – слои
роговицы; 2 – сосудистая оболочка; 2а – пигментный слой сосудистой
оболочки; 3 – радужка; 4 – хрусталик; 5 – задняя камера; 6 – передняя
камера; 7 – кольцевая связка; 8 – сократительная мышца; 9 – серповидный
отросток; 10 – сетчатка; 11 – зрительный нерв
Некоторые особенности строения глаза рыб:
1. Зрачок имеет фиксированный размер (у большинства рыб). Форма может быть круглой, овальной, щелевидной.
2. Хрусталики обычно сферические, изредка – слегка вытянутые, причем более плотные, чем у сухопутных животных.
3. Приближением или удалением хрусталика от сетчатой оболочки с помощью особых мышц могут (в некоторой степени) приспосабливать зрение к расстоянию.
4. Зрение у рыб монокулярное: правый и левый глаз получают отдельное изображение.
Необычна модификация глаз у небольшой рыбы четырехглазки из Центральной и Южной Америки. Глаза у нее помещаются наверху головы, каждый из них разделен перегородкой на две самостоятельные части: верхней частью рыба видит в воздухе, нижней – в воде.
Существует специальный механизм перераспределения палочек, колбочек и пигментных клеток в рыбьем глазу, позволяющий зрению адаптироваться к перемене светового режима. При ярком освещении на передний план выходят колбочки, а в темноте – палочки.
Свет как внешний раздражитель и, следовательно, зрение, имеют неодинаковое значение для разных видов рыб. Планктонофаги и пелагические рыбы значительно зависят от света. При их искусственном ослеплении они утрачивают способность активно питаться. Рыбы-планктонофаги имеют хорошо развитую зрительную систему, у них крупные глаза, большой зрачок.
Для донных рыб (бентософагов) свет и зрение имеют меньшее значение. Так, при ослеплении осетровых их пищевая активность практически не меняется.
Многие хищные рыбы открытых вод при поиске и захвате добычи также полагаются исключительно на зрение, в связи с чем их пищевая активность проявляется только днем. Так, у окуня и судака хорошо развит зрительный анализатор.
Но среди хищных рыб есть и придонные виды, а также виды с пиком активности в ночное время. Понятно, что у этих хищников зрение развито хуже, второстепенно или вообще не имеет значения, по крайней мере, при поиске пиши.
У морских глубоководных рыб, приспособившихся к жизни при очень низкой освещенности, глаза достигают огромных размеров. Диаметр их глаза может составлять от 30 до 50 % от длины головы (Polyipnus spp., Bathymacrops sp., Myctophum spp.). Однако у других глубоководных видов рыб глаза могут быть редуцированы или вообще отсутствовать (Idiacanthus spp., Ipnops spp.) (рис. 45).
Светочувствительный аппарат. Светочувствительный аппарат у рыб включает в себя сетчатку глаз, которая состоит из фоторецепторов двух видов: палочек и колбочек.
Палочки обеспечивают видимость в условиях плохой освещенности, они преобладают в сетчатке ночных и глубоко сумеречных рыб. Колбочки отвечают за остроту зрения и цветоразличение, они преобладают у дневных видов, активные фазы жизни которых проходят в условиях хорошей освещенности.

Рис. 45. Примеры глубоководных
рыб с увеличенными
и редуцированными глазами: А – Myctophum spр., Б – Ipnops spр.
У некоторых представителей рыб, кроме глаз, имеются светочувствительные клетки, разбросанные по поверхности тела.
Основные функциональные параметры зрения у рыб:
1. Угол зрения в вертикальной плоскости составляет до ста пятидесяти градусов, а в горизонтальной – до 170°.
2. Дальность видимости. Большинство рыб способно различать контуры объектов на расстоянии в 10–12 м, это при прозрачной воде и хорошем освещении. Однако рассмотреть детали, т. е. форму, цвет и прочие характеристики, они могут лишь на меньшем расстоянии.
3. Дифференциация объектов. Рыбы могут различать и группировать объекты по внешним признакам. Например, они способны отличить куб от пирамиды или шар от цилиндра.
4. Восприятие движения. Многие активные хищные рыбы имеют хорошую двигательную зрительную реакцию. Мирные рыбы должны издали увидеть приближающуюся опасность, вследствие чего любая подвижность крупных объектов, силуэтов и теней вызывают у них защитную реакцию.
Цветовое зрение. Для большинства рыб характерно цветовое зрение. Однако цвета рыбы воспринимают не в таких красках, как человек. Водная среда может быть сильно пигментирована за счет планктонных организмов или неорганических веществ. Таким образом, вода выступает в качестве светового фильтра. Кроме того, водная поверхность производит поляризацию света, что также приводит к искажению цветовой гаммы.
Цветовое зрение у рыб неодинаково в зависимости от условий среды обитания.
Рыбы, ведущие дневной образ жизни, имеют больше колбочек в сетчатке и лучше различают цвета, чем ночные виды. У рыб, которые живут в мелких и хорошо освещенных местах, есть 4–5 видов колбочек (например, форель, щука, хариус, голавль, язь). В связи с этим они могут различать большое количество цветов.
Другие рыбы, преимущественно ночные донные, имеют два вида колбочек, что ограничивает их возможности. Они тоже различают цвета, но спектр у них гораздо меньше (например, судак, сом, угорь, налим).
Глубоководные рыбы, обитающие на глубине до 1,5 км, различают синий и зеленый цвета. Кроме цвета, рыбы могут различать форму и величину окружающих предметов.
Слух. Установлено, что рыбы слышат и сами издают звуки. Звук представляет собой цепочку регулярно повторяющихся волн сжатия газообразной, жидкой или твердой среды, то есть в водной среде звуковые сигналы так же естественны, как и на суше.
Звукоиздающие органы у рыб различны: плавательный пузырь (это горбыли, губаны), лучи грудных плавников в комбинации с костями плечевого пояса (у сомов), челюстные и глоточные зубы (это окуневые и карповые). В связи с этим неодинаков и характер звуков: они могут напоминать удары, цоканье, свист, ворчанье, хрюканье, писк, кваканье, рычанье, треск, рокот, звон, хрип, гудок. Сила и частота звуков, издаваемых рыбами одного вида, зависит от пола, возраста, пищевой активности, здоровья, и причиняемой боли.
Слуховая система у рыб представлена акустико-латеральной системой. В ее состав входят:
1) внутреннее ухо в костной стенке черепа,
2) плавательный пузырь,
3) веберов аппарат,
4) боковая линия.
Внутреннее ухо (рис. 46) состоит из лабиринта, в который входят преддверие и три расположенных перпендикулярно друг к другу полукружных канала. Внутри перепончатого лабиринта имеется жидкость с отолитами, это слуховые камешки (рис. 47). При воздействии на отолиты звуковых волн они колеблются, и эти колебания воспринимает слуховой нерв.
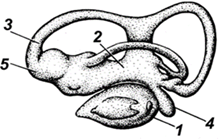
Рис. 46. Орган равновесия и слуха рыб: 1 – круглый
мешочек; 2 – овальный мешочек; 3 – полукружные каналы; 4 –
зачаток улитки; 5 – ампулы
полукружных каналов

Рис. 47. Расположение отолитов во внутреннем ухе клюворылой антиморы
Плавательный пузырь соединен с внутренним ухом, он усиливает вибрации, выполняя функцию резонатора.
Веберов аппарат. У рыб – это орган, соединяющий плавательный пузырь с внутренним ухом у ряда пресноводных костистых рыб (карпообразных, сомовых).
Он состоит из трех пар подвижно сочлененных косточек, которые представляют собой видоизмененные элементы первых трех позвонков (рис. 48).
Функция веберова аппарата – воспринимать, трансформировать в механические смещения и передавать внутреннему уху изменения объема плавательного пузыря – резонатора и преобразователя звука.
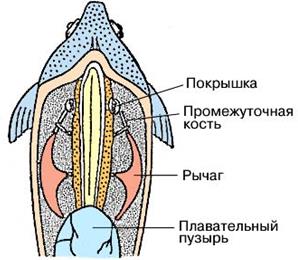
Рис. 48. Схема строения веберова аппарата
Функциональные свойства слуховой системы рыб:
1. Восприятие широкого диапазона частот. Есть разные мнения о том, какие звуки способны воспринимать рыбы. По одним данным, они улавливают частоты от 16 до 60 тыс. Гц, по другим – верхний предел ограничивается 12–13 тыс. Гц. Есть утверждение, что рыбы способны воспринимать весь диапазон звуковых колебаний – от инфра– до ультразвуковых.
2. Выделение важных сигналов. Рыбы способны из всего хаоса окружающих звуков и колебательных проявлений выделять нужные и важные для их жизни сигналы, даже такие слабые, которые находятся на грани возникновения или затухания.
3. Определение направления источника звука. Парные органы слуха рыб стереофонически воспринимают звуковые колебания, что дает возможность четко устанавливать направление и место источника колебания.
4. Реакция на изменения погоды. Возможно, рыбы реагируют на приближение ненастной погоды, благодаря восприятию низкочастотных акустических колебаний, исходящих от зон повышенной конвекции и фронтальных разделов, находящихся вблизи центра циклона.
5. Способность запоминать звуки и связывать их с событиями. Лишний шум на водоеме сильно влияет на поведение рыб, которые имеют хороший слух: они могут перестать передвигаться по водоему в поисках пищи или прервать нерест.
Обоняние. Эта система рыб включает:
Орган обоняния. У большинства видов рыб он хорошо развит и располагается на верхней поверхности головы впереди глаз. Но у эволюционно древних хрящевых рыб, а из костистых – у двоякодышащих, органы обоняния находятся на нижней части головы.
Носовая, или обонятельная полость (носовой мешок). Втянутая внутрь через отверстия – ноздри вода попадает в эту полость, на дне располагаются обонятельные складки, составляющие обонятельную розетку. Поверхность складок покрыта обонятельным эпителием. Дно носовой капсулы выстлано складками эпителия, состоящего из опорных и чувствующих клеток, то есть рецепторов. Наружная поверхность чувствующей клетки снабжена ресничками, а основание связано с окончаниями обонятельного нерва. В обонятельном эпителии многочисленны клетки, вырабатывающие слизь.
Ноздри расположены у хрящевых рыб на нижней стороне рыла впереди рта, у костистых – на дорсальной стороне между ртом и глазами (рис. 49). Круглоротые имеют по одной ноздре, настоящие рыбы – по две. Каждая ноздря разделяется кожистой перегородкой на два отверстия (рис. 50). Вода проникает в переднее из них, омывает полость и выходит через заднее отверстие, омывая и раздражая при этом волоски рецепторов. Под влиянием пахучих веществ в обонятельном эпителии происходят сложные процессы, а именно перемещения липидов, белковомукополисахаридных комплексов и кислой фосфатазы.

Рис. 49. Примеры различного
расположения ноздрей у рыб: 1 – простые,
2 – трубчатые, 3 – свисающие
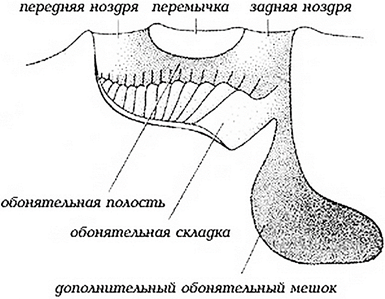
Рис. 50. Схема строения ноздри рыб
Величина ноздрей связана с образом жизни рыб: у подвижных рыб они небольшие, так как при быстром плавании вода в обонятельной полости обновляется быстро; у рыб малоподвижных, наоборот, ноздри большие – они пропускают через носовую полость больший объем воды, что особенно важно для плохих пловцов, в частности обитающих у дна.
Большинство рыб обладают тонким обонянием, то есть пороги обонятельной чувствительности у них очень низки. Это особенно относится к ночным и сумеречным рыбам, а также к живущим в мутных водах, которым зрение мало помогает в поиске пищи и общении с сородичами.
Дополнительные вентиляционные обонятельные мешки предназначены для вентиляции носовой полости и для продукции обонятельной слизи. Рецепторные клетки в таких мешках отсутствуют, но благодаря им через специально развивающееся отверстие может возникать связь между органом обоняния и ротовой полостью.
Первичный обонятельный центр (парные обонятельные луковицы) – это крупный нервный центр, имеющий сложное слоистое строение с большим количеством специализированных нейронов.
Вторичный обонятельный центр – передний мозг. В нем происходит окончательная обработка информации.
Аксоны всех рецепторных клеток после базальной мембраны обонятельного эпителия соединяются в один обонятельный нерв. Он связывает обонятельный орган с первичными обонятельными центрами.
Функциональные свойства обонятельной системы рыб:
1. Высокая чувствительность к запахам. Пороговые концентрации веществ, вызывающие заметные электрофизиологические ответы в обонятельной системе, могут быть крайне низкими.
2. Медленная адаптация. Чувствительность к действующему запаховому стимулу снижается медленно, поэтому не происходит привыкания. Это позволяет рыбам ориентироваться по источнику запаха и перемещаться к нему.
3. Разделение на группы по спектру воспринимаемых запахов. Есть рыбы-макросматики, которые реагируют на широкий спектр запаховых стимулов и проявляют к ним высокий уровень обонятельной чувствительности, т. е. с сильно развитым обонянием, и микросматики, реагирующие только на ограниченный набор запахов, т. е. со слабо развитым обонянием.
4. Роль в поведении. Восприятие обонятельных сигналов позволяет рыбам различать других особей, определять их видовую и популяционную идентичность, а также порождает различные модели поведения: репродуктивные, защитные, миграционные, родительские, территориальные и др.
5. Влияние на работу других органов и систем организма. Обоняние оказывает большое влияние на работу многих органов и систем организма, тонизируя или угнетая их.
6. Зависимость от загрязнения среды. Особенно опасны для обонятельной системы рыб соединения тяжелых металлов и детергенты. Даже очень кратковременное воздействие этих веществ на обонятельную систему приводит к быстрому разрушению обонятельного эпителия и соответственно к полной потере рыбами возможности воспринимать любые запахи.
Осязание. Осязательная система у рыб представлена группами чувствительных клеток, так называемых осязательных телец, расположенных на губах, конце рыла, плавниках и на специальных органах ощупывания – усиках, мясистых выростах. У хрящевых рыб рецепторы осязания расположены на участках тела, не покрытых чешуей, а у костистых – разбросаны по всему телу, но губы и плавники являются наиболее чувствительными. С их помощью рыбы чувствуют прикосновения, боль, температуру воды.
Вкус. Ведущую роль в регуляции заключительной фазы пищевого поведения рыб играет вкусовая рецепция, которая обеспечивает оценку вкусовых свойств пищи и ее соответствие потребностям рыб, а также потребление рыбами адекватных кормовых объектов.
Вкусовые рецепторы у рыб устроены в виде вкусовых почек (вкусовых луковиц), расположеных в ротовой полости и глотке. С их помощью рыбы способны распознавать вкусовые характеристики: соленое, сладкое, кислое, горькое, а также находить подходящую пищу.
Вкусовая почка рыб – это углубление овальной формы, на дне которого расположены группы хеморецепторов, покрытых слизистым слоем. Размеры вкусовых почек у разных рыб колеблются в диапазоне от 30 до 80 мкм по высоте и от 20 до 50 мкм по ширине. Общее число вкусовых почек может достигать у рыб десятков и даже сотен тысяч, что значительно больше, чем у остальных позвоночных животных.
Некоторые особенности строения вкусовой почки рыб
1) основание лежит на базальной мембране эпидермиса.
2) наружу вкусовая почка открывается вкусовой порой, которая может находиться на одном уровне с поверхностью эпидермиса, быть приподнятой или слегка углубленной в эпидермис.
3) в состав входят рецепторные, опорные и базальные клетки, которые окружены полулунными перигеммальными клетками, придающими вкусовой почке характерную форму.
4) рецепторные клетки состоят из трех разных типов, взаимодействующих с разными вкусовыми веществами.
5) базальные клетки располагаются в основании вкусовой почки, и их последующая дифференцировка приводит к образованию всех остальных типов клеток вкусовой почки.
Основания вкусовых почек оплетены концевыми разветвлениями лицевого, блуждающего и языкоглоточного нервов.
У многих видов рыб вкусовые почки располагаются и на внешней поверхности тела – на голове, туловище, плавниках, усиках. Адекватными вкусовыми раздражителями для рыб служат различные типы химических веществ, их смеси и экстракты корма. По уровню вкусовой чувствительности рыбы значительно превосходят многих других позвоночных животных. У одной и той же особи количество вкусовых почек растет по мере увеличения размеров тела.
У рыб, питающихся в светлое время суток, у хищников, полагающихся на зрение при активном преследовании своих жертв, наружные вкусовые почки малочисленны либо полностью отсутствуют. У рыб с сумеречным или ночным пиком питания и у бентосоядных рыб наружные вкусовые почки покрывают большую часть тела или всю его поверхность.
Чувствительность органов вкуса у некоторых рыб очень высока. Так, пещерные рыбы семейства Харациновых (Anoptichthys jordani), будучи слепыми, ощущают раствор глюкозы в концентрации 0,005 %.
Боковая линия – это совокупность специализированных кожных органов чувств, расположенных правильными рядами по бокам туловища и на голове рыб (рис. 51).
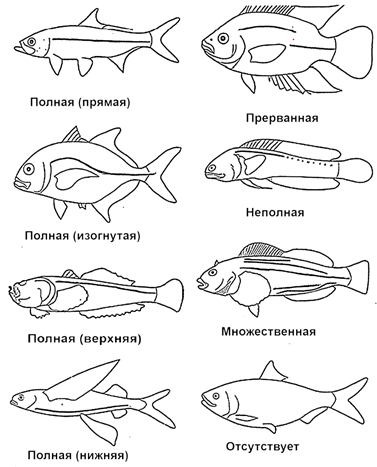
Рис. 51. Различные типы боковых линий у рыб
Структурная организация: у большинства рыб органы боковой линии лежат под кожей в тянущихся вдоль тела каналах, которые сообщаются с внешней средой отверстиями в чешуйках (рис. 52). Основной механорецепторной единицей органов боковой линии служит невромаст, содержащий чувствительные волосковые клетки. Они похожи на волосковые клетки внутреннего уха. Это говорит о том, что эти органы имеют общее происхождение.
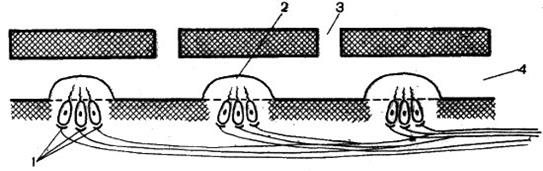
Рис. 52. Схема
строения боковой линии: 1 – волосковые клетки
(нейромасты); 2 – купула; 3 – пора; 4 – канал
Иннервация: на рецепторных клетках невромастов оканчиваются разветвления нервных волокон. Чувствительные клетки боковой линии оканчиваются на вершине сосочками или волосками, а с противоположной стороны – веточкой нерва. Смещение сосочка или волоска создает генераторный потенциал, который передает информацию по нервам в акустико-латеральный центр мозга.
· ориентируются в пространстве. Могут ориентироваться даже в условиях полной темноты, знают рельеф и характер дна;
· определяют направление и скорость движения воды;
· распознают членов стаи. Помнят характерные для «одногруппников» инфразвуковые звуки;
· регулируют пищевое поведение. Улавливают движение мотыля на дне и знают не только место его расположения, но и предположительное количество;
· обнаруживают приближение врага или жертвы. Впереди плывущей рыбы бегут волны, отражаясь от подводных предметов, и, возвратившись к рыбе, воспринимаются ее боковой линией;
· координированно перемещаются в стае.
Вестибулярная система рыб. Эта система предоставляет информацию о движении головы и пространственной ориентации, относительно силы тяжести. Она обеспечивает стабильность взгляда, равновесие и контроль позы рыбы.
Вестибулярная система рыб представлена внутренним ухом – органом слуха и равновесия, который расположен в задней части черепной коробки. Таким образом, в одном органе объединены слуховая функция и функция равновесия. Ушных отверстий и ушных раковин у рыб нет.
В эпителии, выстилающем внутреннюю часть ушного лабиринта, находятся чувствительные клетки с волосками. Полости лабиринта заполнены жидкостью, которая называется эндо-лимфа.
При движении рыбы отолиты (рис. 53) смещаются в ту или иную сторону и давят на поля волосковых клеток, благодаря чему рыба может определять свое положение в пространстве и воспринимать звуковые колебания. Кроме того, меняется давление эндолимфы (вязкой жидкости, которая создает среду для восприятия звуковых волн и движений головы, обеспечивает питание и вымывание продуктов обмена веществ из рецепторных клеток, поддерживает электрохимический потенциал, необходимый для передачи нервных импульсов) в полукружных каналах, и возникшее раздражение регистрируется нервными окончаниями.
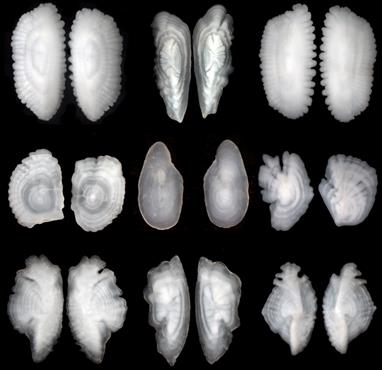
Рис. 53. Отолиты различных видов рыб
Рыбы воспринимают низкочастотные звуки боковой линией, а высокочастотные – внутренним ухом.
Электрорецепция – способность обнаруживать электрические поля или токи. Некоторые рыбы, обладающие электрорецепцией:
– акулы имеют органы электрорецепторов, которые позволяют обнаруживать электромагнитные поля, создаваемые всеми живыми существами (рис. 54), что помогает им находить добычу;
– нильский слоник – использует электрорецепцию для навигации в мутных водах (рис. 55).
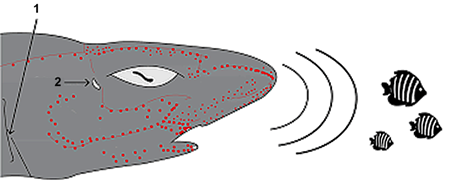
Рис. 54. Схема электрорецепции у
акул: 1 – жаберное отверстие,
2 – брызгальце
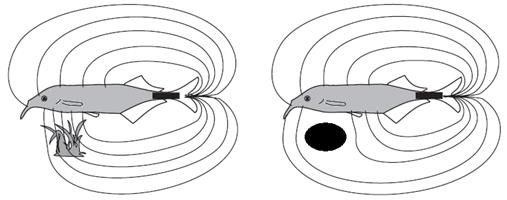
Рис. 55. Схема электрорецепции у нильского слоника
Роль электрорецепции в поведении рыб:
1. Ориентация в пространстве. Например, нильский слоник Gnathonemus petersii из семейства мормировых, который обитает в центральной и западной Африке, в покрытых густой растительностью участках рек с небольшим течением и мутной, темной водой, излучает электрический сигнал, а рецепторы на его теле реагируют на отражение от предмета, благодаря чему рыба понимает, как далеко от нее он находится.
2. Общение с сородичами. С помощью электрических полей рыбы обмениваются различной информацией, которая может быть агрессивно-оборонительной, групповой, межполовой и опознавательной. Например, африканские рыбки из отряда мормирид используют электрические импульсы для призыва партнеров для спаривания.
3. Поиск пищи. Акулы, скаты и некоторые пресноводные рыбы (сом, щука) обладают высокой чувствительностью к внешним электрическим полям и различают на некотором расстоянии движущихся рыб по их биопотенциалам, то есть выделяемым биоэлектрическим сигналам.
Электрочувствительные рецепторы рыб делятся на два типа: ампулярные и бугорковые (рис. 56). У некоторых рыб присутствуют оба типа рецепторов, а у большинства – только один.
![]()
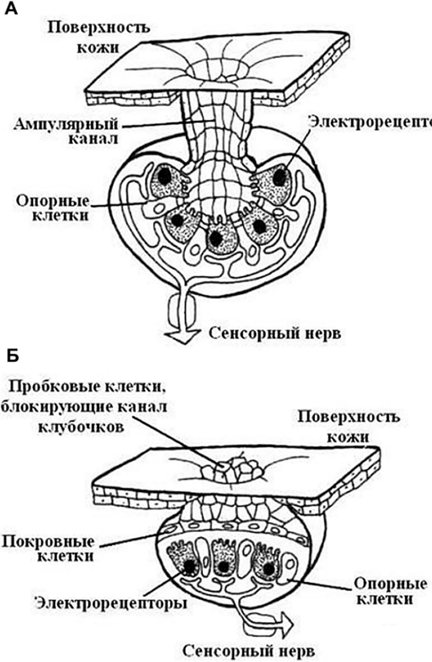
Рис. 56. Различные типы
электрочувствительных рецепторов у рыб:
А –
ампулярные, Б – бугорковые
Структурная организация: электрические сигналы, достигнув рецепторных волосковых клеток ампулы или клубочка, преобразуются в рецепторный потенциал волосковой клетки, откуда сигнал передается на окончание сенсорного нейрона.
У акул и скатов электрорецепторы расположены на голове. Это ампулы Лоренцини. У нильского слоника электрорецепторы находятся в нижней губе, внешне напоминающей хобот.
Функциональные свойства:
– ампулярные электрорецепторы приспособлены для восприятия медленно изменяющихся электрических полей;
– бугорковые рецепторы реагируют на быстрые изменения. Они менее чувствительны, чем ампулярные.
Электрорецепторы позволяют рыбам чувствовать электрическое поле и любые его изменения, вызванные окружающим миром или другими рыбами. Это помогает им ориентироваться в пространстве, различать формы и размеры объектов, получать сигналы от других рыб.
Хеморецепция – способность воспринимать химические вещества и реагировать на их изменения через свое поведение.
Рыбы имеют три крупные хемосенсорные системы (рис. 57): обонятельную, вкусовую и общей химической чувствительности. Каждая из них обладает хорошо развитым рецепторным аппаратом и мозговыми центрами.
С помощью хеморецепции рыбы:
– определяют состав среды обитания;
– оценивают качество пищи;
– опознают сородичей, отличают своего от чужака, врага от мирного соседа;
– разыскивают пищу, находят дорогу к местам нереста, откорма.
По способу хеморецепции вкус и осязание осуществляются тактильно, контактно, обоняние – дистанционно.
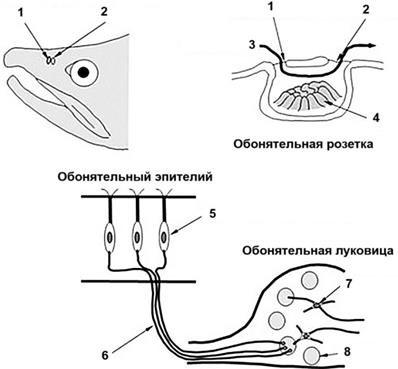
Рис. 57. Схема обонятельной
системы рыб: 1 – передняя ноздря; 2 – задняя ноздря; 3 –
поток воды; 4 – обонятельная пластинка; 5 – обонятельный
рецепторный нейрон; 6 – обонятельный нерв; 7 – митральная клетка;
8 – клубочки
Механорецепция – это восприятие механических раздражителей из окружающей среды – воды, подводных объектов, растений, камней, пищевых организмов и контактов с сородичами. Для механорецепции у рыб используются различные рецепторы.
1) узкоспециализированные механорецепторы – тактильные нейроны и свободные нервные окончания, которые разбросаны по всему телу и расположены или между эпителиальными клетками или под ними. Специализированные механорецепторы различного строения, то есть луковицы, спирали, диски, концентрируются на коже головы, плавниках, усиках;
2) механорецепторы сухожилий – при помощи них локомоторные центры спинного мозга рыб осуществляют обратную связь с мышцами, то есть оценивают степень их укорочения и напряжения.
3) внутренние органы – сильные механические воздействия на тело рыбы отражаются на состоянии внутренних органов, например плавательного пузыря, который имеет развитую систему рецепторов.
Магниторецепция – способность чувствовать магнитные поля Земли, которая обнаружена у некоторых животных, в т. ч. у рыб (рис. 58).
Рыбы, например лососи, используют магниторецепцию для ориентации во время миграций на большие расстояния через океаны и реки. Эта способность позволяет им находить свои места нереста даже после долгих лет путешествий в открытом море.
Ученые предполагают, что магниторецепция может быть связана с присутствием магнитита – минерала, чувствительного к магнитным полям, который находится в тканях рыб. Если рыбы оказываются в зоне магнитных аномалий, их чутье может дать сбой, и они теряют ориентацию в пространстве.
Рис. 58. Отряды рыб, у
представителей которых отмечено наличие
магниторецепции (синий цвет) и в отношении которых исследования наличия
магниторецепции пока не проводились (оранжевый цвет)
Магниторецепция играет важную роль в жизни рыб, позволяя им ориентироваться в пространстве и осуществлять навигацию.
Роль магниторецепции в жизни рыб
1. Миграция. Рыбы используют магнитное поле Земли не только для поддержания постоянного курса, но и для определения местоположения в море и возвращения в родовые районы. Например, лососи применяют геомагнитные сигналы, чтобы вернуться к месту своего рождения. Считается, что молодь лосося, скатываясь из родной реки в океан, запоминает свойства магнитного поля в ее окрестностях. Эти данные выступают в качестве географического ориентира, который рыбы используют для возвращения на нерестилище, когда станут взрослыми.
2. Ориентация. В присутствии геомагнитного поля рыбы выбирают преимущественно направление запад – восток или юго-юго-запад – северо-северо-восток.
3. Двигательная активность. Магнитные поля влияют на двигательную активность рыб: в однородном постоянном поле, например, угри предпочитают двигаться вдоль линий индукции, а в градиентном – в направлении уменьшения интенсивности.
4. Синхронизация биологических часов. Колебания магнитного поля могут служить синхронизатором биологических часов, давая возможность рыбам отмечать ход времени.
Способность воспринимать магнитные поля есть у рыб разного эволюционного уровня. Пластиножаберные рыбы могут воспринимать поля с помощью ампулярных органов, а у некоторых костистых рыб для магнито-рецепции служат рецепторные клетки, содержащие цепочки кристаллов биогенного магнетита.
Вопросы для самоконтроля
1. Какие пять отделов головного мозга есть у рыб?
2. Где расположен спинной мозг рыб? Его функции?
3. Какие органы чувств есть у рыб?
4. Какие пять особенностей есть в строении глаза рыб?
5. Какие пять функциональных свойств слуховой системы есть у рыб?
6. Где расположены ноздри у хрящевых и у костистых рыб? Как величина ноздрей связана с образом жизни рыб?
7. Как устроен вкусовой рецептор у рыб? (Краткое описание). Где расположены эти рецепторы?
8. Какие шесть видов деятельности выполняют рыбы с помощью боковой линии?
9. Что такое хеморецепция? Какие четыре вида деятельности выполняют рыбы с ее помощью?
10. Что такое магниторецепция? Какова роль магниторецепции (четыре фактора) в жизни рыб?