ОТДЕЛ ЗЕЛЕНЫЕ ВОДОРОСЛИ – CHLOROPHYTA.
Общая характеристика. Особенности цитологической организации
Размножение. Циклы развития. Принципы деления на классы.
Класс Вольвоксовые.
Класс Протококковые или Хлорококковые.
Класс Улотриксовые.
Класс Коньюгаты, или Сцеплянки.
.
Общая характеристика. Особенности цитологической организации.
Представители отдела характеризуются травяно-зеленой окраской хлоропластов сходной с окраской высших растений. У них обнаружены хлорофиллы а и b, а и b - каротиноиды и около 10 различных ксантофиллов. Запасное вещество - крахмал. Обитают зеленые водоросли преимущественно в пресных водах, где, по-видимому, возникли и прошли основные этапы своей эволюции, хотя имеются и морские виды. В морфологическом отношении зеленые водоросли отличаются многообразием по сравнению с другими отделами, у них представлены все известные ступени морфологической дифференциации тела водорослей, кроме амебоидной и тканевой. Основные структуры тела следующие:
- Монадная, характеризуется наличием у клеток жгутиков 2, 4 и более, способных к активному движению. Форма клетки - продолговато-овальная, нередко грушевидная, шаровидная. Одноклеточные, ценобиальные и колониальные организмы. У простейших водорослей монадная структура наблюдается в течение всей их вегетативной жизни, а у высокоорганизованных водорослей лишь в клетках, служащих для бесполового (зооспоры) или полового (гаметы) размножения.
- Пальмеллоидная или капсальная структура - соединения нескольких или многих неподвижных клеток, погруженных в общую слизь временно или постоянно.
- Коккоидная - неподвижные клетки, имеющие оболочки различной формы, одиночные или соединенные в колонии разнообразного типа, но не нитчатые.
- Нитчатая структура - сложение неподвижных клеток в нити, состоящие из одного или нескольких рядов клеток.
- Гетеротрихальная (разнонитчатая). Водоросли состоят из стелющихся по субстрату нитей горизонтальных и отходящих вверх от них вертикальных.
- Сифональная или сифоновая организация отличается отсутствием клеточных перегородок, так что талломы, часто крупных размеров, имеющие внешнюю расчлененность, формально - одна клетка с большим количеством ядер. Неклеточное строение.
- Сифонокладальная - многоядерные клетки, соединенные в нитчатые или иной формы многоклеточные талломы.
Клетки многих примитивных, имеющих монадную организацию, водорослей, а также зооспоры и гаметы большинства водорослей «голые», т.е. ограничены снаружи только цитоплазматической мембраной (плазмолеммой). У большинства водорослей кнаружи от плазмолеммы находится клеточная стенка, состоящая из целлюлозы и пектиновых веществ. Нередко в толще клеточной стенки присутствуют добавочные компоненты: кремний (педиаструм), карбонат кальция (харовые водоросли). У некоторых водорослей в стенке имеется хитин (кладофора). Клеточная стенка может нести различные выросты.
В цитоплазме различимы элементы ЭПС, рибосомы, митохондрии, аппарат Гольджи, ядро или ядра, хроматофоры (хлоропласты). Хроматофоры различны по форме, многочисленные или одиночные. В матриксе хроматофоров находятся включения - пиреноиды, особые белковые тельца, вокруг которых откладывается крахмал или близкие к нему углеводы.
Некоторое своеобразие имеет цитологическая организация водорослей монадной структуры. У них нет клеточной стенки. Цитоплазма занимает всю клетку. Нет вакуолей с клеточным соком, характерных для питательных растительных клеток. Имеющиеся вакуоли - пульсирующие, выполняют экскреторную функцию, выделяют из клетки ненужные продукты обмена веществ. Монадные клетки имеют кирпично-красный глазок или стигму, состоящий из двух частей - передний неокрашенный, двояковыпуклый, аналогично хрусталику глаза, и задний, более или менее чашевид-ный, содержащий пигмент близкий к каротину. Глазок - орган зрения, ориентирующий движение клетки под влиянием световых лучей (фототаксис). Монадные водоросли имеют жгутики, от одного до многих, преобладают двухжгутиковые формы (изоморфные или гетероморфные).
Размножение. Циклы развития. Принципы деления на классы.
Обычно различают вегетативное, бесполое и половое размножение.
Вегетативное размножение - процессы размножения, при которых части таллома отделяются без каких-либо заметных изменений в протопласте. Это деление клеток, у колониальных - распад колоний или образование новых поколений внутри клеток материнской особи. Более специализированная форма вегетативного размножения - образование толстостенных, переполненных запасными продуктами клеток, которые предназначены для перенесения неблагоприятных условий, их называют акинетами.
Бесполое размножение осуществляется посредством особых спор, покидающих материнскую клетку. Из содержимого каждой клетки может сформироваться только одна зооспора (эдогониум) или 2, 4, 8 и более зооспор. Зооспоры имеют монадную структуру, не одеты оболочкой. У некоторых водорослей вместо зооспор образуются аплапоспоры или автоспоры неподвижные, без жгутиков. Гемизооспоры или амебоиды - наиболее примитивные споры бесполого размножения. От зооспор они отличаются отсутствием жгутиков, а от автоспор, кроме того, и отсутствием настоящей оболочки, в связи с чем способны производить амебоидные движения. У ряда групп водорослей бесполое размножение отсутствует.
Половой процесс у водорослей состоит в слиянии двух специализированных гаплоидных клеток, называемых гаметами в одну диплоидную клетку, называемою зиготой.
Хологамия - самый примитивный тип полового процесса - сливаются две взрослые особи (некоторые вольвоксовые).
Изогамия - сливаются подвижные и одинаковые по форме и величине гаметы.
Гетерогамия или анизогамия - сливаются подвижные гаметы, разных размеров. Большая крупная - женская, маленькая - мужская. Клетки, в которых образуются изо- и гетерогаметы - гаметангии.
Оогамия - высший тип полового процесса. В этом случае мужская подвижная маленькая гамета - сперматозоид оплодотворяет более крупную неподвижную яйцеклетку. Клетки, в которых формируются сперматозоиды, называются антеридиями, а клетки, содержащие яйцеклетки, оогонии.
Коньюгация - сливается содержимое двух вегетативных недифференцированных клеток, выполняющих функцию гамет.
Растения, производящие гаметы, могут быть обоеполыми - гомоталличными и раздельнополыми - гетероталличными.
В результате полового процесса образуется зигота. Чаще всего зигота окружается толстой оболочкой с запасом питательных веществ. Прорастает зигота сразу или более или менее продолжительного периода покоя.
Соотношение диплоидной и гаплоидной фаз в жизненном цикле развития водорослей неодинаково. В одних случаях прорастание зиготы сопровождается редукционным делением, при этом развивающиеся растения оказываются гаплоидными, и, единственная диплоидная стадиия в цикле - зигота, а вся вегетативная фаза проходит у них в гаплоидном состоянии, они являются гаплобионтами. У других водорослей, наоборот, вся вегетативная фаза диплоидна, гаплоидная фаза представлена гамета-ми, перед образованием которых и происходит редукционное деление ядра. Зигота без редукционного деления прорастает в диплоидный таллом. Растения - диплобионты. У третьих, редукционное деление ядра предшествует образованию зооспор и аплапоспор, развивающихся на диплоидных талломах. Зооспоры прорастают в гаплоидные растения, размножающиеся только половым путем. После слияния гамет зигота развивается в диплоидное растение, несущее только органы бесполого размножения. У этих водорослей имеет место чередование поколений: диплоидного бесполого спорофита и гаплоидного полового гаметофита. Оба поколения могут быть одинаковы морфологически (изоморфная смена поколений) или резко различны по внешнему виду (гетероморфная смена поколений).
Классифицируются зеленые водоросли до сих пор по-разному, и устоявшаяся общепринятая система отсутствует. Во многих системах отдел зеленых водорослей делят на 5 классов:
Кл. Вольвоксовые – клетки со жгутиками, талломы монадные, ценобиальные или колониальные
- Кл. Протококковые – клетки без жгутиков, талломы коккоидные или колониальные
- Кл. Улотриксовые – многоклеточные нитчатые и пластинчатые формы.
- Кл. Сифоновые – неклеточные одноядерные или многоядерные формы с различным внешним видом.
Класс Вольвоксовые –Volvocaphyceae.
Сюда относятся монадные, т.е. снабженные жгутиками, одноклеточные, ценобиальные и колониальные водоросли, подвижные в течение вегетационной жизни. Жгутиков, чаще 2 (1 - 3). Ядро шаровидное, в центре клетки, хлоропласт один, чашевидный париентальный с одним крупным пиреноидом. В передней части глазок, есть пульсирующие вакуоли. Большинство автотрофы, известны гетеротрофы и миксотрофы. Размножение вегетативное, бесполое и половое (разнообразные формы). Зигота прорастает после периода покоя.
Вольвоксовые - типичные обитатели планктона. Обитают преимущественно в мелких, стоячих, нередко быстро пересыхающих водоемах. Это активные санитары загрязненных и сточных вод. Их используют в качестве показательных форм при биологическом анализе загрязненных вод.
Среди представителей: дюналиелла солевая, вызывающая «красное цветение» воды в перенасыщенных солью водоемах - одноклеточная водоросль, покрытая тонким и нежным перипластом (нет клеточной стенки). В процессе жизнедеятельности в ней накапливаются каротиноиды, придающие окраску. Размножение у нее в основном вегетативное, путем продольного деления клетки в подвижном состоянии. Половой процесс – хологамия.
Порядок хламидомонадовые - Chlamydomonadales.
Семейство хламидомонадовые - Chlamydomonadaceae
Род хламидомонада (500 видов) - Chlamydomonas
Представителями семейства являются картерия, хлорогониум, гематококк и хламидомонада. Хламидомонады – это активные санитары загрязненных вод, в которых они бурно размножаются, вызывая зеленое зацветание. Клетки способны через оболочку поглощать органические вещества, что способствует процессам самоочищения водоема. Это значит, что наряду с автотрофным питанием хламидомонаде свойственно и гетеротрофное.
Это одноклеточная водоросль эллипсоидальной формы с заметным выпячиванием, впереди, которая несет два одинаковых жгутика (рис.10).
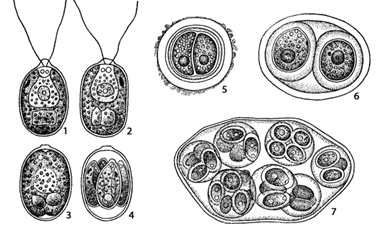
Рис. 10. Chlamydomonas: 1 – внешний вид клетки;
2–4 – процесс образования зооспор; 5–7 – пальмеллевидное состояние и размножение внутри слизи
Оболочка у некоторых представителей может быть мягкой, легко растягивающейся, или очень плотной. Протопласт содержит одно гаплоидное ядро в углублении чашевидного хроматофора. Хроматофор имеет один пиреноид в крахмальной сфере. А вверху, на его поверхности – и глазок. В передней части клетки находятся две сократительные вакуоли. При подсыхании водоемов хламидомонады переходят в пальмелоидное состояние. При этом клетки теряют жгутики, стенка их ослизняется и происходит размножение. Дочерние клетки не расходятся, они тоже претерпевают деление, а их клеточные стенки, в свою очередь ослизняются.
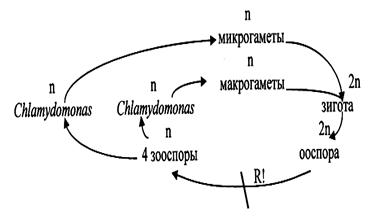
В благоприятных условиях размножение бесполое (с помощью, зооспор). При этом клетки останавливаются и последовательно в продольном направлении делятся на 2, 4 или 8 частей. Эти дочерние клетки вырабатывают жгутики, и после ослизнения материнской оболочки выплывают наружу в виде мелких зооспор, которые растут до размера материнской особи. Половой процесс изогамный (рис.11). Он происходит при неблагоприятных условиях. Гаметы образуются внутри материнской клетки так же, как и зооспоры, но в большем количестве и более мелких размеров. Т.о., особь является гаметоспорофитом. Описаны гетерогамия и оогамия, но это исключения. У многих видов наблюдается гетероталлизм (сливаются гаметы различных разно заряженных особей). После слияния ядер сливаются и пластиды. Вначале зигота сохраняет жгутики и плавает. Затем, жгутики сбрасываются, и выделяется плотная оболочка (первичная и чуть позже вторичная). Диплоидная зигота впадает в состояние покоя. В течение этого периода она или остается прежних размеров, или увеличивается. Но в любом случае в ней накапливаются питательные вещества, поэтому многие ботаники считают зиготу самостоятельно существующим организмом - спорофитом. Затем зигота прорастает. Ее содержимое при этом делится мейотически с образованием четырех зооспор. Таким образом, все стадии жизненного цикла у хламидомонады (за исключением зиготы) протекают в гаплоидном состоянии. Таким образом, цикл развития хламидомонады антитетический (со сменой поколений): гаплоидный одноклеточный гаметофит, способный к половому и бесполому размножению и диплоидный одноклеточный спорофит (так называемая зигота)
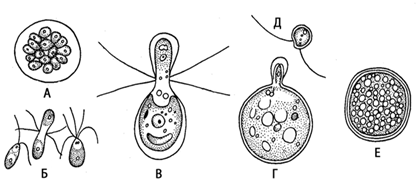
Рис. 11. Половой процеес у Chlamydomonas:
А – образование изогамет; Б – изогамия;
В – гетерогамия; Г – оогамия; Д – мужская гамета; Е – зигота
Цикл развития Chlamydomonas
Порядок вольвоксовые - Volvocales
Семейство вольвоксовые Volvocaceae
Род вольвокс Volvox
Вольвокс – обитатель пресноводных водоемов. Это шаровидная колония до 5 мм в диаметре (рис.12,А). Клетки изодиаметричные со жгутиками, связаны между собой плазмодесмами и расположены одним слоем по поверхности. Внутренняя полость шара заполнена жидкой слизью. Клетки разнотипные.
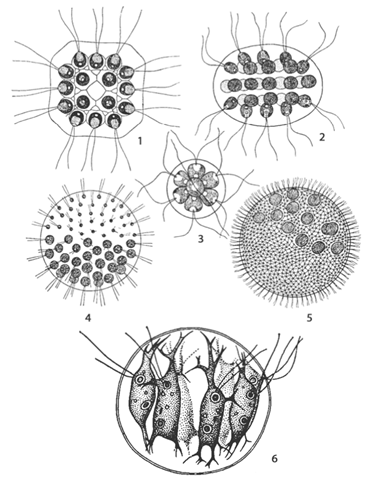
Рис. 12. Представители порядка Volvocales:1 – Gonium; 2 – Eudorina; 3 – Pandorina; 4 – Pleodorina; 5 – Volvox; 6 – Stephanosphaera
Имеются вегетативные клетки. Есть партеногонидии (гонидии), которые способны давать, дочерние колонии. Есть клетки – антеридии (дают двужгутиковые сперматозоиды) и клетки – оогонии (содержат одну яйцеклетку). Т.о., половой процесс оогамный. Образовавшаяся зигота переходит в состояние покоя, после чего ее диплоидное ядро редукционно делится с образованием четырех зооспор: В каждой из них сформируется колония в результате бесполого размножения. При этом протопласт претерпевает столько делений, чтобы сформировать полный комплекс клеток будущей колонии. Вначале в результате клеточных делений из клеток сформируется пластинка (гоническая стадия), затем из нее образуется полая сфера с маленьким незамкнувшимся отверстием. На этой стадии жгутики находятся внутри, т.е., направлены к центру сферы. В дальнейшем незамкнутая сфера полностью выворачивается наизнанку, жгутики приобретают нормальную ориентацию, а отверстие сферы полностью замыкается. Маленькая колония растет до положенного размера. Таким же образом формируются дочерние колонии в партеногонидиях.
Цикл развития Volvox

Класс Протококковые –Protococcophyceae.
Объединяет коккоидные формы, в основном это одноклеточные и ценобиальные формы, реже колонии. В эволюционном плане зеленых водорослей на этом этапе впервые возникла, утвердилась и приобрела широкое развитие коккоидная, типично растительная структура тела. По строению протопласта протококковые напоминают вольвоксовых. Наиболее примитивные из них сохранили пульсирующие вакуоли, глазок и даже жгутики, хотя последние неподвижные и называются псевдоцилиями. Наличие этих рудиментальных органелл доказывает происхождение протококковых от вольвоксовых. Громадное большинство - микроскопические формы, лишь у немногих родов таллом может достигать крупных размеров. Форма клеток разнообразна, но преобладает шаровидная, элипсовидная и яйцевидная. Оболочка клеток всегда сплошная, из целлюлозы, реже с примесью пектиновых веществ, у низших форм - пектиновая. У многих видов она снаружи снабжена щетинками, иглами, шипиками, бородавками или слизью, что помогает клетке парить в толще воды.
Основная масса протококковых водорослей - планктонные формы. Для них характерна миксотрофность, некоторые виды встречаются в симбиозе с другими организмами. Есть среди протококковых виды - эндофиты, живущие в инфузориях, в лишайниках.
У протококковых водорослей преобладает бесполое размножение, реже наблюдается половой процесс, иногда они размножаются путем простого деления клеток. Бесполое размножение осуществляется гемизооспорами, зооспорами и автоспорами. Половой процесс встречается редко, в основном, это - изогамия, еще реже - гетерогамия. Оогамия известна лишь у немногих родов.
Порядок хлорококковые – Chlorococcales
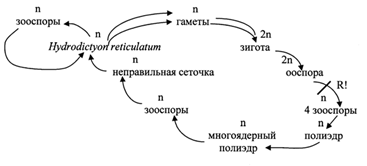
Семейство гидродикциевые - Hydrodictyaceae
Род гидродикцион - Hydrodictyone
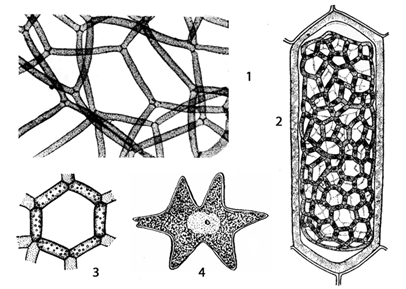
Рис. 13. Hydrodictyon: 1 – участок взрослого ценобия; 2 – молодая сеточка внутри материнской клетки; 3 – часть ювенильного ценобия; 4 – полиэдр
При бесполом размножении протопласт материнской клетки раскалывается на множество участков, которые превращаются в одноядерные двужгутиковые зооспоры (20 000). Некоторое время они движутся внутри оболочки материнской клетки, а затем втягивают жгутики, выделяют собственную оболочку и соединяются, образуя новую сеточку, которая освобождается после разрыва материнской оболочки. Молодая сеточка растет лишь за счет увеличения слагающих ее клеток, причем число ядер в них увеличивается.
Половой процесс изогамный. Любая материнская клетка при этом раскалывается на большое число (30 000) мельчайших изогамет. Через отверстие в материнской клетки они выплывают в воду, где и сливаются между собой. Для гидродикциона характерна гомоталличность. У некоторых видов копулируют гаметы даже одной клетки. Образовавшаяся зигота покрывается плотной оболочкой и переходит в состояние покоя. Позже, в благоприятных условиях, она прорастает. При этом ее ядро редукционно делится и образуется четыре гаплоидные двужгутиковые зооспоры. Они плавают некоторое время в воде, затем останавливаются и каждая развивается в многоугольную клетку - полиэдр. Он разрастается, становится многоядерным и его содержимое распадается на зооспоры, которые слагаются в молодую зародышевую сеточку, освобождающуюся через разрыв в оболочке полиэдра.
Цикл развития Hydrodictyon reticulatum
Класс Улотриксовые – Ulothrichophyceae.
К улотриксовым относятся водоросли, имеющие нитчатое строение, а также некоторые пластинчатые и мешковидные, которые все же в начале онтогенеза имеют нитчатую форму. Все клетки растения могут участвовать в росте растения, все могут образовывать споры и гаметы, за исключением клеток основания нити, которыми они прикрепляются. Клетки тела автономны: способны к регенерации, вегетативному размножению, а также способны становиться репродуктивными.
При бесполом размножении во всех зеленых клетках развивается от 2 до 16 (32) четырехжгутиковых зооспор. Обычно этот процесс начинается в апикальной (конечной клетке) и продвигается к основанию нити. После периода движения зооспора останавливается, сбрасывая один за другим жгутики, прикрепляется к субстрату и прорастает в нить. При половом размножении в клетках возникают двухжгутиковые гаметы в числе 4 (8) - 32 (64). Половой процесс чаще изогамный, преобладает гетероталлизм. Четырехжгутиковая зигота прорастает в одноклеточный спорофит (спорофит впадает в период покоя), который при созревании распадается на 4 - 16 четырехжгутиковых зооспор. Зооспоры ведут себя так же как и зооспоры бесполого поколения.
Порядок улотриксовые - Ulothrichales
Семейство улотриксовые - Ulothrichaceae
Род улотрикс - Ulothrix
Вид улотрикс опоясанный - Ulothrix zonata
Обитает в быстротекущих ручьях или прибойной полосе пресноводных озер. Слоевище состоит из однорядных неветвящихся нитей неопределенной длинны. Все клетки одинаковы, за исключением базальной, которая вытягивается в короткий ризоид и прикрепляет нить к каменистому субстрату, и верхушечной, которая куполообразно закруглена. Клетка улотрикса имеет толстую оболочку, крупную центральную вакуоль, постенную цитоплазму с одним гаплоидным ядром и одним пластинчатым хроматофором в виде незамкнутого кольца. В хроматофоре содержится несколько пиреноидов. (рис14).
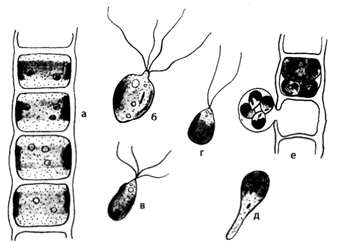
Рис. 14. Ulothrix zonata: а – внешний вид нити; б, в – зооспоры; г – гамета; д – прорастание зооспоры; е – образование зооспор
Все клетки (за исключением базальной) способны делиться в поперечном направлении. Это способствует непрерывному диффузному нарастанию таллома, а также размножению организма.
Цикл развития Ulothrix zonata
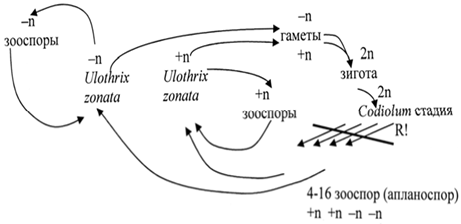
Любая клетка гаплоидной нити, за исключением базальной, может производить 2 – 32 четырех жгутиковые зооспоры. Из них через некоторое время после контакта с субстратом вырастают новые нити. Та же нить (дающая зооспоры) в ухудшающихся условиях существования образует двужгутиковые гаметы. Половой процесс - изогамия. Зигота вырабатывает толстую оболочку и впадает в период покоя. Перед ее прорастанием происходит мейоз и образуется от 4 до 16 гаплоидных зооспор (иногда неподвижных апланоспор), так же прорастающих в нити. Поэтому данный цикл развития считали гаплонтным, а сам организм – гаметоспорофитом. Однако у морских обитателей этого же самого вида из зооспор зиготы образуются одноклеточные длительно существующие (весь вегетативный сезон) спорофиты. В этом случае жизненный цикл является антитетическим с гетероморфной сменой поколений, где преобладает гаметофит (нить).
Порядок ульвовые - Ulvales
Семейство ульвовые - Ulvaceae
Род ульва – Ulva
Усложнение изоморфного цикла развития наблюдается у представителей порядка ульвовые. Ульва (морской салат) это макроскопическая морская водоросль теплых мелководий (рис.15).
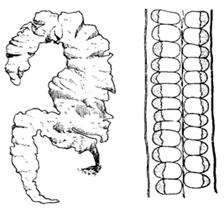
Рис15. Внешний вид (слева) и поперечный срез (справа) таллома Ulva
Таллом ее имеет вид сильно гофрированной по краям пластинки паренхимного строения. В нижней части имеется основание с ризоидными отростками, с помощью которых осуществляется прикрепление к субстрату. Клетки способны делиться в двух и более плоскостях.
Четырехжгутиковые зооспоры и двужгутиковые гаметы никогда не формируются на одном и том же экземпляре. Одни растения бесполые (диплоидные спорофиты) и образуют в результате редукционного деление гаплоидные зооспоры; другие - половые (гаплоидные гаметофиты), вырастающие из зооспор. Они продуцируют только изогаметы. После копуляции изогамет диплоидная зигота прорастает сразу, без периода покоя и без мейоза в спорофит. Гаметангии и спорангии у ульвы не дифференцированы, поэтому гаметы и зооспоры образуются в обычных клетках. Тип развития антитетический со сменой изоморфных поколений (спорофит и гаметофит внешне неотличимы).
Класс Коньюгаты, или Сцеплянки – Conjugatophyceae.
Коньюгаты - одноклеточные и многоклеточные нитчатые водоросли. Космополиты, встречаются на всех континентах. Живут неприкрепленными в пресных водоемах, реже в сырых местах в земле. Класс включает около 4500 видов. Их особенности - полное отсутствие жгутиковых стадий (зооспор и снабженных жгутиками гамет) и особый тип полового процесса в виде коньюгации. Образование зигот у коньюгат - явление довольно редкое. В вегетативном состоянии все гаплобионты. В большинстве случаев одноклеточные коньюгаты размножаются обычным вегетативным делением в поперечной плоскости, а нитчатые - распадением нитей на отдельные клетки. Клетки коньюгат одноядерные, хроматофоры крупные, лентовидные.
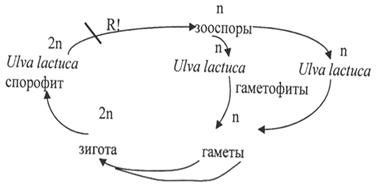
Своебразный половой процесс - коньюгация.
При половом размножении обычно две нити располагаются параллельно друг другу. Супротивные клетки образуют выросты, направленные друг к другу и срастающиеся концами. Оболочки их в месте соприкосновения растворяются, и получает-ся сквозной канал, через который содержимое одной клетки в течении нескольких ми-нут перемещается в другую и сливается с ее протопластом, сжавшимся к этому времени. Чаще содержимое клеток соединяется в коньюгационном канале. Кроме описан-ной выше лестничной коньюгации изредка наблюдается боковая коньюгация (иногда у тех же видов) - через боковой канал нах поперечной перегородкой содержимое клетки переходит в соседнюю клетку той же нити.
У одноклеточных водорослей при половом размножении две сблизившиеся клетки окружаются слизью, оболочки клеток раздвигаются и выходящие из них протопласты сливаются.
Зигота прорастает после периода покоя. Этому предшествует редукционное деление диплоидного ядра.
В основе деления класса на порядки: морфологический тип тела, особенности клеточной оболочки.
Порядок зигнемовые - Zygnematales
Семейство спирогировые - Spirogyraceae
Род спирогира - Spirogyra
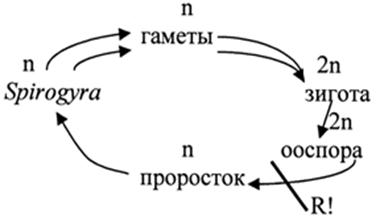
Спирогира (340 видов) это пресноводная нитчатая однорядная неветвящаяся водоросль. Могут иметься ризоиды. Нити переплетены между собой и окружены общей слизью. В длину она растет за счет поперечных делений ее клеток, которые имеют вытянутую цилиндрическую форму. Все клетки абсолютно одинаковые с целлюлозно-пектиновой клеточной стенкой. Характерны хроматофоры в виде одной или многих лент, расположенных в постенной цитоплазме и опоясывающие клетку по спирали. По средней линии хроматофоров расположены пиреноиды, окруженные крахмальными зернами. Гаплоидное ядро одно и расположено оно в центре клетки в цитоплазматическом мешочке, подвешенном на цитоплазматических тяжах, отходящих от постенной цитоплазмы в центре вакуоли.
Конъюгация может быть лестничной и боковой (рис.16). Наиболее обычна последняя, которая происходит между клетками двух нитей. Нити располагаются параллельно друг другу и вначале склеиваются слизью. Затем противолежащие клетки образуют навстречу друг другу выросты, которые соприкасаются и срастаются своими концами.
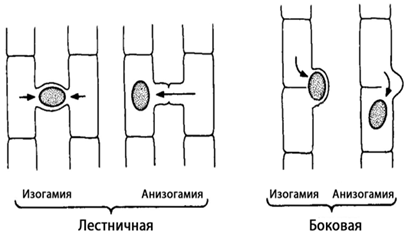
Рис. 16. Схема конъюгации у зигнемовых
Удлиняясь, выросты постепенно раздвигают нити, и возникает фигура в виде лестницы. Стенки на концах соприкасающихся отростков растворяются, и возникает узкий конъюгационный канал. Протопласт одной из противолежащих клеток (отдающей) сокращается, отделяется от стенок и переползает в другую (воспринимающую) клетку, где сливается с ее содержимым, образуя зиготу. У некоторых видов слияние протопластов и образование зиготы происходит в конъюгационном канале. Образовавшаяся зигота округляется, выделяет толстую трехслойную оболочку и переходит в состояние покоя. При прорастании происходит редукционное деление. Из четырех гаплоидных ядер остается жизнеспособным только одно, которое и разовьется в гаплоидную нить. Цикл развития спирогиры происходит по гаплонтному типу, особь является гаметофитом.
Цикл развития Spirogyra
Порядок десмидиевые -Desmidiales
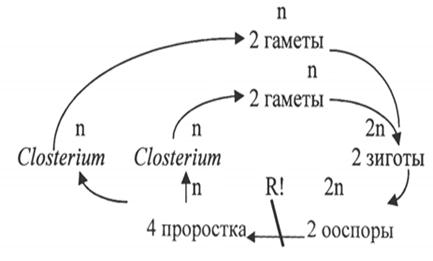
Включает одноклеточные (клостериум, космариум, микрастериас), реже нитчатые (десмидиум, гиалотека) гаплоидные водоросли. (рис.17). Их клетки всегда состоят из двух симметричных половинок – полуклеток. В плоскости симметрии имеется заметная перетяжка. Полуклетки могут быть различной формы. Клеточная стенка состоит из двух половинок, находящих друг на друга в плоскости симметрии, она целлюлозосодержащая. Наружная поверхность (химическая природа не установлена) может нести шипики, бородавочки и другие выросты.
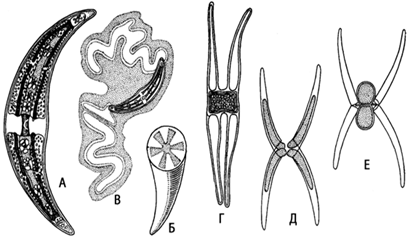
Рис. 17. Closterium: А – вид клетки сбоку; Б – поперечный разрез клетки; В – выделение масс слизи с концов клетки; Г – конъюгация;
Д, Е – конъюгация с образованием двойных зигот
Кроме того, на ней имеются поры, выделяющие слизь, за счет чего клетки осуществляют свое движение. Ядро одно и расположено оно в области перешейка полуклеток. Хроматофоры осевые или постенные с пиреноидами. Они в форме пластинок или лент. В клетке их может быть от одного до нескольких. Вакуоли небольшие и могут располагаться по концам клетки. Размножение вегетативное и половое (конъюгация). При конъюгации клетки сближаются и покрываются общей слизью. Между ними образуется канал, внутри которого сливаются протопласты. У многих десмидиевых половинки оболочек конъюгирующих клеток расходятся, протопласты освобождаются и сливаются без образования конъюгационного канала. При вегетативном размножении ядро делится надвое и каждое отходит в одну из полуполовинок. В середине перешейка образуется перегородка. Каждая полуклетка строит новую свою половину и лишь после этого, достигнув положенного размера, они отделяются.
Цикл развития Closterium