ОТДЕЛ ДИАТОМОВЫЕ ВОДОРОСЛИ – BACILLARIOPHYTA.
Общая характеристика. Особенности химического состава и строения клеточных стенок. Размножение.
Класс Центрические.
Класс Перистые, или Пеннатные.
Значение диатомовых водорослей.
Общая характеристика. Особенности химического состава и строения клеточных стенок. Размножение.
Диатомовые водоросли - это особая группа одноклеточных микроскопических буровато-желтых организмов, одиночно живущих или объединенных в колонии различного типа: цепочки, нити, ленты, звездочки, кустики. Они широко распространены в бентосе и планктоне морей (центрические диатомеи) и пресных (пеннатные диатомеи) водоемов. Кроме того, они поселяются в болотах, моховых подушках, на камнях, скалах, почвах, на поверхности льдов и снегов, придавая им бурую окраску. Наиболее разнообразны по видовому составу и количеству диатомеи ценозы обрастания (в том числе днища кораблей, кожа китов и других животных). Многие водоросли этого отдела являются индикаторами солености, приуроченные к определенным ее величинам. Однако есть виды, степень чувствительности которых к NaCL не так высока.
Резким отличием диатомеи от других водорослей является структура и состав их оболочки, покрывающей клетки снаружи. Оболочка пропитана кремнеземом (от 4 до 50 %) и называется панцирем. Целлюлоза в ней отсутствует. Стенки панциря пронизаны регулярно повторяющимися мельчайшими отверстиями (ареолами), с помощью которых обеспечивается обмен между протопластом и внешней средой. Характер расположения этих отверстий и их количество специфичны для разных родов и видов. Форма оболочки, ее толщина и скульптурная поверхность (гребни, складки, шипики, выпуклости, щетинки, пустоты, поры) очень разнообразны и также специфически для разных таксонов. По форме панциря все диатомеи делятся на две группы: центрические (радиально-симметричный панцирь) и пеннатные или перистые (двусторонне-симметричный панцирь). Панцирь вырабатывается самой клеткой. Он состоит из двух частей и по конструкции напоминает коробку, закрытую крышкой. Большая часть панциря (крышка) называется эпитекой. Она своими краями находит на внутреннюю (меньшую), которая называется гипотекой. Эпитека и гипотека состоят из створки и поискового ободка (поясок). (рис.20).

Рис.20. Строение и форма панциря пеннатных диатомовых водорослей.
Слева: 1 – вид со створки; 2 – вид с пояска; 3 – поперечный разрез клетки (э – эпитека, г – гипотека, с1 – створка эпитеки, с2 – створка гипотеки,
по1 – поясковый ободок эпитеки, по2 – поясковый ободок гипотеки, п – поясок, зс1 – загиб створки эпитеки, зс2 – загиб створки гипотеки, оп – осевое поле, цп – центральное поле, ш – шов (ш1 – шов эпитеки,
ш2 – шов гипотеки), цу – центральный узелок, ку – конечный узелок, р – ребра).
Справа вверху: 1 – овальная; 2 – таблитчато-прямоугольная; 3 – линейная;
4 – ланцетная; 5 – ромбическая; 6 – гитаровидная; 7 – S-образная;
8 – полулунная; 9 – дуговидная; 10 – скобковидная; 11 – ладьевидная; 12 – яйцевидная; 13 – булавовидная; 14 – клиновидная. Справа внизу:
половина панциря Grammatophora со вставочными ободками (вс) и септами (с)
Именно створки придают клеткам очертания, которые могут быть округлыми, эллиптическими, палочковидными, ромбическими, треугольными, серповидными, клиновидными. Поясок эпитеки налегает на поясок гипотеки, плотно его охватывает, но не срастается с ним. У многих диатомей образуются еще вставочные ободки, от одного до многих, отличающихся не только формой, но и структурой. Благодаря им происходит увеличение роста и объема клетки. Большинство диатомеи перистого типа характеризуются присутствием шва. Это сквозная щель (или две щели), прорезывающая стенки створки от ее концов к середине. У отдельных представителей шов может находиться всего на одной створке или даже лишь на ее половинке; у других – на обеих створках. Швы соединяются с помощью узелков в трех местах: в середине и у концов. Биологическое значение шва очень велико, т.к. через него происходят не только обменные процессы с внешней средой, но и осуществляется быстрое передвижение по субстрату и в толще воды. Это происходит благодаря трению движущейся цитоплазмы о воду или субстрат или благодаря выделяющейся через шов слизи.
Цитоплазма в клетках диатомеи располагается постенным слоем, или скапливается в центре клетки и у ее полюсов. Остальное пространство заполнено множеством вакуолей, которые иногда сливаются в одну крупную. Ядро диплоидное и располагается в цитоплазматическом мостике или в периферическом слое цитоплазмы. Хроматофоры у большинства центрических диатомеи мелкие, многочисленные в форме зерен или дисков без пиреноидов. У пеннатных водорослей хроматофоры обычно крупные, пластинчатые, занимают почти всю полость клетки и с пиреноидами. Они имеют желтую или желто-бурую окраску, что зависит от пигментов, среди которых преобладают бурые – каротин, ксантофилл (в том числе фукоксантин) и диатомин, маскирующие в живой клетке хлорофиллы а и с. После гибели бурые пигменты растворяются в воде и зеленый хлорофилл становится видимым. Состав и количество пигмента непостоянно, а зависит от интенсивности света, химического состава воды и возраста водоросли. Некоторые диатомовые обладают способностью переходить от автотрофного питания к гетеротрофному. Известны также облигатные гетеротрофы. Их хроматофоры или бесцветные, или их нет совсем. В качестве запасных питательных веществ выступают капли масла, волютин, хризоламинарин и лейкозин.
Чаще всего у диатомовых водорослей происходит вегетативное размножение делением клетки на две половинки. Этот процесс своеобразен из-за наличия твердого панциря. Сначала протопласт увеличивается в объеме из-за скапливающегося масла. Вследствие этого эпитека и гипотека расходятся. После этого протопласт делится параллельно створкам на две равные части. Все органоиды до деления распределяются равномерно между половинками. Если хроматофор один, то он делится пополам. После окончательного разделения клетки на две дочерние каждая из них получает лишь одну половинку панциря. А вторую (обязательно меньшую – гипотеку) достраивает сама. Т.о. возникшие две дочерние клетки по размерам оказываются несходными. Одна (получившая материнскую эпитеку) сохраняет размеры материнской клетки. Другая (получившая материнскую гипотеку) приобретает меньшие размеры. Поэтому после многократных делений размеры клеток у половины особей постоянно уменьшаются. Но этому противостоит половой процесс.
Бесполое размножение не изучено, т.к. природа образующихся микроспор не установлена, поэтому для диатомей характерен диплонтный тип цикла развития с гаметической редукцией.
Половое размножение диатомовых водорослей всегда связано с ауксообразованием, не свойственное другим водорослям. Встречаются все типы полового процесса: изогамия, гетерогамия, оогамия. Кроме того, имеет место автогамия и конъюгация. Оплодотворению у диатомовых предшествует редукционное деление ядра, поскольку вегетативные клетки диплоидные.
У пеннатных диатомей половой процесс – конъюгация изоморфных безжгутиковых гамет. Две особи сближаются и выделяют слизь. В каждой клетке ядро редукционно делится с образованием четырех гаплоидных. У одних видов дегенерирует три ядра, у других два. В первом случае весь протопласт становится гаметой во втором - образуется две гаметы. При копуляции гамета одной клетки, двигаясь амебоидно, переползает к гамете другой клетки. В результате образуется одна или две зиготы, которые сразу, без стадии покоя быстро увеличиваются в размерах и превращаются в ауксоспору. При этом тонкая оболочка зиготы разрывается, а зрелые ауксоспоры вырабатывают кремневую оболочку, превращаясь в вегетативную клетку. Конъюгирующие клетки, лежащие далеко друг от друга образуют копуляционные каналы для прохождения гамет. Протопласты обеих клеток могут выходить в канал и сливаются попарно.
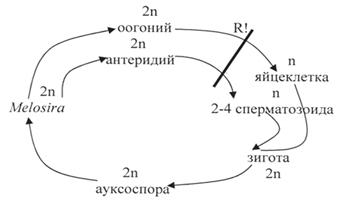
У центрических водорослей – оогамный половой процесс. При этом в одних клетках образуется четыре сперматозоида с одним или двумя жгутиками. В других клетках три ядра дегенерируют, а оставшиеся становится яйцеклеткой. Свободноплавающие сперматозоиды оплодотворяют ее.
У определенных представителей при половом процессе в обеих клетках жизнеспособны все ядра, которые превращаются в изогаметы. В результате образуются четыре зиготы. У Melosira и Chaetoceros происходит автогамия. В этом случае два из четырех гаплоидных ядер дегенерируют, а два оставшихся сливаются друг с другом с восстановлением диплоидного ядра.
В современной системе диатомовые водоросли делят на два класса: центрические и пеннатные.
Класс Центрические – Centrophyceae.
Створки имеют радиальную симметрию, через них можно провести не менее трех плоскостей симметрии. Клетки одиночные или соединены в нитевидные или цепочковидные колонии. Форма панциря различная. Створки в очертаниях круглые, эллиптические, треугольные и многоугольные; их поверхность плоская или выпуклая, иногда вогнутая, реже волнистая. Шва нет. По краю створки расположены выступы в виде выростов, шипов, щетинок, при помощи которых клетки соединяются в колонии. Хлоропласты в виде многочисленных мелких зерен или одной или нескольких пластинок. Различные виды центрических диатомей широко представлены в планктоне Мирового океана, а также пресных континентальных водоемов – рек, озер, водохранилищ, где они являются одними из главных продуцентов органического вещества. Это очень древняя группа, представители известны еще с раннего мела; к настоящему времени многие роды целиком вымерли, из других родов сохранились только некоторые виды. Класс Centrophyceae делится на ряд порядков, из них наиболее крупные – Thalassiosirales, Melosirales, Coscinodiscales.
В планктоне морей широко распространены виды рода косцинодискус (Coscinodiscus) (рис. 21, 1). Клетки внешними очертаниями имеют сходство с цилиндром, диаметр которого часто меньше высоты. На створках радиальными или тангентальными (расположенными по хорде окружности панциря) рядами, реже в беспорядке располагаются ареолы, по краю створки имеются шипики.

Рис. 21. Центрические диатомовые водоросли: 1 – Coscinodiscus (панцирь со створки); 2 – Cyclotella (а – вид со стороны створки, б – с поясковой стороны); 3 – Melosira; 4 – Chaetoceros (в – клетка со створки, г – часть колонии)
В морских и пресных водах, иногда на дне водоемов встречаются виды рода циклотелла (Cyclotella) (рис. 21, 2) либо в виде одиночных клеток, либо, реже, в виде колоний из рыхлых цепочек. Краевая зона створки несет радиальные штрихи или ребрышки, центр створки часто более выпуклый, у большинства видов бесструктурный. Хлоропласты мелкие, пластинчатые, расположены в периферическом слое цитоплазмы.
Цилиндрическую или боченковидную форму имеют панцири видов рода мелозира (Melosira) (рис. 21, 3), обитающих в планктоне и бентосе пресных и соленых водоемов. Клетки соединены в плотные нити (колонии) своими створками при помощи слизи, иногда в скреплении створок принимают участие и шипики. Створки круглые, покрыты неж- ными гранулами. Створки соединяются своими расширенными загиба- ми в середине клетки. Поясковые ободки появляются в период деления клетки. Загиб створки большой, покрыт беспорядочно расположенными мелкими гранулами. Хлоропласты многочисленные в виде пластинок, дисков или лопастные. Ниже представлен цикл развития центрической диатомовой водоросли на примере Melosira.
Цикл развития Melosira
У видов рода хетоцерос (Chaetoceros) (рис. 21, 4) формируются це- почковидные колонии, в которые объединяются клетки с помощью длинных полых щетинок или шипов, расположенных на краях створок. У ряда видов образуются вставочные ободки, увеличивающие объем клетки.
Класс Перистые, или Пеннатные - Pennatophyceae.
Водоросли одноклеточные или колониальные. Со стороны пояска створки имеют двустороннюю симметрию, через них можно провести не более двух плоскостей симметрии. Панцирь симметричный по продольной оси, изопольный, реже гетеропольный, иногда дорсовентральный, редко асимметричный, с пояска линейный, таблитчатый, клиновидный, прямой, иногда S-образно изогнутый, часто с вставочными ободками и септами. Створки линейные, ланцетные, эллиптические, булавовидные, серповидные, овальные, яйцевидные и очень редко круглые, симметричные по продольной оси, изопольные или гетеропольные, реже симметричные по поперечной оси . По продольной оси створки проходит узкая бесструктурная полоса – осевое поле; в середине створки осевое поле часто расширяется, образуя центральное поле. У большинства видов вдоль середины осевого поля расположен щелевидный шов. Иногда шов сдвинут к краю створки или находится в киле (каналовидный шов). Хлоропласты в виде одной или нескольких крупных пластинок, реже – в виде многочисленных мелких пластинок.
Пеннатные диатомеи – пресноводные и морские водоросли, обитающие в бентосе на различных субстратах, и только единичные виды являются планктонными. Класс пеннатных диатомей моложе центрических.
Порядок Araphales – Бесшовные. Клетки одиночные или соединены в пучки, звездчатые или зигзаговидные колонии, в вегетативном состоянии неподвижны. Панцирь прямой. Створки от эллиптических до линейных, иногда булавовидные. Осевое поле от нитевидного до широ- колинейного, без шва.
У видов рода синедра (Synedra) (рис. 22, 1) клетки одиночные или соединены в пучковидные колонии. Панцирь с пояска палочковидный. Створки прямые, линейные, к концам суженные, с одним пластинчатым хлоропластом и тонкими, нежными параллельными штрихами, прерванными осевым полем. Со стороны пояска клетка имеет вид сильно вытянутого прямоугольника с двумя ребрами хлоропластов, тонкими, нежными штрихами, расположенными по загибу створки. Встречается в морских, солоноватых и пресных водоемах, в бентосе и обрастаниях, редко в планктоне.
Часто встречающиеся в пресноводных и морских водах представите- ли рода астерионелла (Asterionella) (рис. 22, 2) – типичные компоненты планктона. Клетки соединены в звездчатые колонии (aster – звезда). Панцирь с пояска линейный, гетеропольный (базальный конец шире головного). Створки узколинейные, с головчатыми концами, головка на базальном конце крупнее, чем на свободном конце. Виды рода фрагилярия (Fragilaria) (рис. 22, 3) напоминают Synedra, но соединяются створками при помощи шипиков в длинные лентовид- ные или загзагообразные колонии, обитающие в прибрежной зоне озер, прудов и других водоемов, среди высших водных растений, в обрастаниях, реже в планктоне. С пояска панцирь узкий, линейный. Форма створок от узколинейной до ланцетной, они часто расширены посередине, иногда волнистые или треугольные.
Род табеллярия (Tabellaria) (рис. 22, 4) объединяет широко распространенные пресноводные виды, часто встречающиеся в планктоне и бентосе различных водоемов с мягкой маломинерализованной водой с повышенной цветностью. Клетки соединены в звездчатые, лентовидные или зигзаговидные цепочки. Клетки с пояска имеют вид табличек, снабжены вставочными ободками и септами, створки линейные или овальные, на концах или на середине расширенные.

Рис. 22. Диатомовые бесшовные водоросли: 1 – Synedra (а – панцирь с пояска; б, в – со стороны створки); 2 – Asterionella (г – панцирь со створки;
д – общий вид колонии); 3 – Fragilaria (е – панцирь с пояска; ж, з – со створки); 4 – Tabellaria (и – общий вид колонии; к – панцирь со створки)
Порядок Raphales – Шовные. Это типичные обитатели бентоса пресных и морских водоемов, а также множество видов приурочено к сообществам обрастаний разных предметов, погруженных в воду, и высших водных растений. Клетки большей частью одиночные, подвижные, реже собраны в лентовидные или кустовидные колонии. Панцирь продольно или поперечно симметричный, створки от линейных до широко эллиптических, изредка S-образно изогнутые. Одна или обе створки имеют шов. Структура створки представлена штрихами, ребрами, которые располагаются поперечными рядами, иногда камерами.
Род пиннулярия (Pinnularia) (рис. 23, А–Д) – наиболее известный среди шовных диатомовых водорослей. Клетки одиночные, редко соединенные в ленты. Панцирь с пояска прямоугольный. Створки изопольные, линейные, линейно-эллиптические и ланцетные, с закругленными концами. Структура створок состоит из гладких поперечных ребер, представляющих собой полые камеры в толще панциря. Шов нитевидный, центральный и конечные узелки хорошо развиты. Хлоропласты в виде двух пластинок, расположенных вдоль поясковых сторон клетки. Населяет преимущественно пресные водоемы, реже моря, часть видов – ископаемые. Со стороны створки видны ребра двух хлоропластов, а со стороны пояска – один пластинчатый. Смещение хлоропластов со створковой стороны на поясковую связано с появлением шва.
Виды рода навикула (Navicula) (рис. 23, Е) имеют много общего с видами Pinnularia, но отличаются от последней отсутствием камер в створках. Структура створки состоит из нескольких поперечных ребрышек, состоящих из ареол разной формы (округлых, эллипсоидных). Концы клеток более заостренные, суженные, по форме створки напоминают лодочку (navicula по-латыни – лодочка).
Виды гиросигмы (Gyrosigma) (рис. 23, Ж) имеют створки, по форме напоминающие букву S и имеющие штриховку панциря в виде мелкой сеточки.
Из водорослей с каналовидным швом очень разнообразны в бентосе, обрастаниях и реже в планктоне водоемов виды рода нитцшия (Nitzschia) (рис. 23, И). Створки имеют вид палочек с заостренными концами. На каждой створке имеется особая продольная складка – киль с каналовидным швом, идущим вдоль каждой створки клетки.

Рис. 23. Шовные диатомовые водоросли: Pinnularia: А – клетка со створки; Б – клетка с пояска;
В – деление клетки (две дочерние клетки с пояска); Г – панцирь со створки; Д – панцирь с пояска; Е – Navicula (панцирь со створки); Ж – Gyrosigma
(панцирь со створки); З – Cymbella (а – панцирь со створки, б – колония); И – Nitzschia (панцирь со створки).
1 – шов; 2 – центральный и конечный узелки; 3 – ядро;
4 – капли масла; 5 – хлоропласт; 6 – цитоплазматический мостик
Значение диатомовых водорослей.
Их называют пастбищами морей, т.к. их продуктивность велика, а содержание белков и жиров у них выше, чем в картофеле и хлебных злаках. Поэтому они являются мощным источником органического вещества и служат первоначальным звеном в пищевых цепях для многих организмов: сельдь, хамса, сардины и др. И в отмершем виде они идут на питание бактериям и простейшим, а также участвуют в процессе осадконакопления, образуя диатомовые илы. Известны породы под названием диатомит (горная мука, кизельгур), состоящие на 50 – 80 % из панцирей диатомовых водорослей. Они очень легкие, пористые и рыхлые. Применяются как полировочный и шлифовальный материал, для тепловой и звуковой изоляции, как фильтрующее вещество в пищевой, химической и медицинской промышленности. По ископаемым формам можно определить происхождение и возраст различных осадочных пород. Диатомеи деятельно участвуют в процессах естественного очищения воды. Есть виды индикаторы, которые используют при оценке санитарного состояния прибрежных вод. Однако массовое развитие некоторых диатомовых водорослей может иметь и отрицательные последствия, т.к. они (Thalassiosira decipiens) в больших количествах попадают в жабры личинок рыб и моллюсков и вызывают их гибель.