ЛЕКЦИЯ 1.
ОБЩАЯ ХАРАКТЕРИСТИКА ГРИБОВ
Грибы - особое царство организмов.
Особенности строения клетки. Строение вегетативного тела.
Половой процесс и способы размножения, типы спороношений.
Ядерные фазы в цикле развития грибов.
Принципы классификации и систематика грибов.
Грибы особое царство организмов
Царство грибов характеризуется следующими признаками:
- образование хорошо выраженной клеточной стенки;
- абсорбтивное питание;
- размножение спорами;
- неподвижность в вегетативном состоянии и неограниченный рост;
- первично гетеротрофный способ питания;
- запасной продукт - гликоген.
Грибов известно около 100000 видов. В царстве грибов различают следующие основные классы:
Класс хитридиомицеты - Chytridiomycctes.
Класс гифохитриомицеты - Hyphochytriomycetcs.
Класс оомицеты - Oomycetes.
Класс зигомицеты - Zygomycetes.
Класс аскомицеты - Ascomycetes.
Класс базидиомицеты - Basidiomycetes.
Класс дейтеромицеты, или несовершенные грибы - Deuteromycetes.
В современных классификациях грибы выделяют как самостоятельное царство организмов, коренным образом, отличающееся и от растений и от животных.
Выделение грибов в качестве самостоятельного царства косвенно подтверждается данными по сравнительному изучению цитохромов C у разных групп организмов. Эти исследования показывают, что грибы - древняя группа, существовавшая еще до расхождения растений и животных.
Особенности строения клетки. Строение вегетативного тела
Клетка большинства грибов имеет хорошо выраженную клеточную стенку, состоящую до 80-90% из полисахаридов, связанных с белками и липидами. Кроме того, в ее состав входят полифосфаты, пигменты, например меланины, и другие вещества. Микрофибриллярные скелетные компоненты клеточной стенки состоят из хитина или целлюлозы.
В цитоплазме клеток грибов хорошо различимы рибосомы, митохондрии, аппарат Гольджи и ядра (рис. 1). Протопласт грибов окружен цитоплазматической мембраной - плазмалеммой. Между клеточной стенкой и цитоплазматической мембраной грибов располагаются ломасомы - мембранные структуры, имеющие вид многочисленных пузырьков. В зависимости от происхождения различают настоящие ломасомы и плазмалеммасомы. Последние представляют собой производное плазмалеммы.
На границе между цитоплазмой и вакуолью также расположена мембрана - тонопласт. Между тонопластом и плазмалеммой находится внутренняя мембранная система - эндоплазматическая сеть.
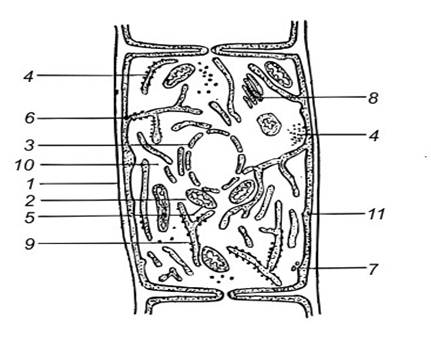
Рис. 1. Схема строения грибной клетки:
1 - клеточная стенка, 2,5 - митохондрии, 3 - ядро, 4 - рибосомы, 6,11 - цитоплазматическая мембрана, 7 - ломасомы, 8 - аппарат Гольджи (диктиосомы), 9- эндоплазматическая сеть, 10 - цитоплазма
Митохондрии грибов сходны с митохондриями растений, но отличаются от них некоторыми деталями строения. Строение крист митохондрий у разных групп грибов неодинаково. Кристы оомицетов и гифохитриомицетов имеют трубчатое строение, сходное с кристами высших растений. У остальных групп грибов кристы митохондрий пластинчатые.
В клетках грибов находится от одного до 20-30 ядер. Их размер обычно около 2-3 мкм. Ядра грибов имеют типичное строение. Они окружены оболочкой, состоящей из двух мембран. В нуклеоплазме содержатся ядрышко и хромосомы. При митотическом делении ядра ядерная оболочка часто сохраняется.
Подвижные клетки грибов - зооспоры и гаметы - имеют жгутики. Жгутики грибов построены по типу, характерному для всех эукариот. Их главный стержень состоит из двух одинарных центральных микротрубочек, окруженных кольцом из девяти дублетов микротрубочек. Жгутики у грибов двух типов: бичевидные гладкие и перистые. У перистых жгутиков по бокам располагаются два ряда мастигонем - полых трубочек 2-3 мкм длиной и около. 20 нм диаметром. Крупные таксоны грибов, представители которых имеют жгутики, различаются по их количеству и расположению на клетке.
При традиционном делении живых организмов на царства животных и растений грибы относили к растениям. Хотя грибы существенно отличаются от растений неспособностью к фотосинтезу и соответственно гетеротрофным способом питания, их объединяли на основе таких черт сходства, как хорошо выраженная клеточная стенка, абсорбция питательных веществ из растворов, отсутствие большей частью подвижности в вегетативном состоянии. Однако гетеротрофный способ питания накладывает отпечаток на характер обмена у грибов. По таким признакам, как присутствие в обмене мочевины, образование в качестве запасного продукта гликогена, а не крахмала, а также содержание хитина грибы сходны с животными. Большинство грибов отличается от растений и по такому признаку, как путь синтеза лизина. Если у всех растений синтез этой аминокислоты идет через диаминопимелиновую кислоту, у большинства грибов он происходит при участии аминоадипиновой кислоты.
Вегетативное тело большинства грибов представляет собой мицелий, состоящий из ветвящихся нитей – гиф с апикальным (верхушечным) ростом и боковым ветвлением. Мицелий пронизывает субстрат и всей поверхностью поглощает из него питательные вещества (субстратный мицелий), а также располагается на его поверхности и может подниматься над субстратом (поверхностный и воздушный мицелий). На воздушном мицелии обычно образуются органы размножения.
Различают неклеточный, или ценоцитический мицелий, лишенный перегородок и представляющий как бы одну гигантскую клетку с большим числом ядер (рис. 2, Б), и клеточный, или септированный мицелий, разделенный перегородками - септами на отдельные клетки, содержащие от одного до многих ядер (рис. 2, В). Для представителей классов хитридиомицетов, оомицетов, гифохитриомицетов и зигомицетов, условно называемых низшими грибами, характерен неклеточный мицелий. У всех высших грибов - аскомицетов, базидиомицетов и дейтеромицетов - мицелий клеточный, с настоящими септами, образующимися упорядоченно, синхронно делению ядер. Септы иногда находят и у низших грибов: они отделяют репродуктивные органы и поврежденные участки гиф.

Рис. 2. Вегетативное тело грибов.
А - одноклеточный таллом с ризомицелием; Б - неклеточный мицелий; В - клеточный мицелий; Г- почкующиеся клетки
У некоторых грибов, например дрожжей, вегетативное тело представлено одиночными почкующимися или делящимися клетками (рис. 2, Г). Если такие почкующиеся клетки не расходятся, образуется псевдомицелии.
Некоторые примитивные грибы имеют одноклеточный таллом, иногда лишенный клеточной стенки. Одноклеточные талломы грибов часто развивают ризомицелий - разветвленные нитевидные структуры, лишенные ядер (рис. 2, А).
При формировании плодовых тел и некоторых вегетативных структур гифы грибов плотно переплетаются, образуя ложную ткань - плектенхиму. По происхождению она отличается от настоящей ткани, возникающей в результате деления клеток в трех направлениях. Настоящие ткани у грибов встречаются довольно редко. Тканевое строение имеют, например, талломы лабульбениевых - Laboulbeniales из класса аскомицеты.
У многих грибов гифы соединяются параллельно в мицелиальные тяжи, выполняющие проводящие функции. Хорошо развитые и дифференцированные мицелиальные тяжи называют ризоморфами. Они известны, например, у опенка осеннего. Ризоморфы нередко достигают нескольких метров длины. Гифы их наружных слоев имеют утолщенные, часто темноокрашенные стенки и выполняют защитную функцию, а внутренние гифы - проводящую.
Другой тип видоизменений мицелия представляют широко распространенные у многих групп грибов склероции - плотные переплетения мицелия, служащие для перенесения неблагоприятных условий. Клетки склероциев богаты запасными питательными веществами. Часто склероции дифференцированы на кору - наружные слои клеток, обычно толстостенных и темноокрашенных, и внутреннюю часть, состоящую из тонкостенных светлоокрашенных клеток.
Половой процесс и способы размножения, типы спороношений
Грибы размножаются вегетативным, бесполым и половым путем. При вегетативном размножении от мицелия отделяются неспециализированные его части, которые дают начало новому мицелию. Одной из форм вегетативного размножения можно считать и образование хламидоспор - толстостенных клеток, предназначенных для перенесения неблагоприятных условий. У дрожжей вегетативное размножение происходит путем почкования клеток.
Бесполое размножение происходит при помощи специализированных клеток (реже многоклеточных структур) - спор. Споры у грибов развиваются эндогенно – внутри спорангиев или экзогенно - на специализированных веточках мицелия – конидиеносцах (рис. 3, В).
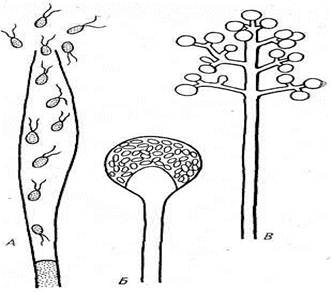
Рис. 3. Органы бесполого размножения грибов.
А - зооспорангий; Б - спорангий; В - конидиеносец с конидиями
У некоторых грибов бесполое размножение происходит при помощи зооспор - голых подвижных клеток, снабженных жгутиками. Зооспоры образуются эндогенно - в зооспорангиях (рис. 3, А). У зигомицетов в спорангиях развиваются неподвижные споры, называемые спорангиоспорами. Спорангии находятся на специализированных гифах - спорангиеносцах, поднимающихся над субстратом (рис. 3, Б). Конидии, как и спорангиоспоры,- неподвижные споры, пассивно переносимые токами воздуха, каплями дождя, насекомыми и другими способами. Однако в отличие от эндогенных спорангиоспор они образуются на мицелии или его специализированных ветвях – конидиеносцах экзогенно.
Известно несколько типов формирования конидий из конидиогенных клеток. Конидиеносцы грибов имеют весьма разнообразное строение, часто наблюдается их агрегация в коремии, ложа или образование полузамкнутых структур - пикнид, в которых развиваются конидиеносцы и конидии. Бесполое размножение при помощи конидий характерно для аскомицетов, дейтеромицетов, базидиомицетов и некоторых низших грибов.
Половое размножение отмечено у всех групп грибов, кроме дейтеромицетов. Формы полового процесса у грибов разнообразны (рис. 4). Их можно разделить на три большие группы: гаметогамия, гаметангиогамия и соматогамия.
Гаметогамия может быть:
- изогамной - слияние морфологически не различающихся гамет;
- гетерогамной - слияние гамет, различающихся по размерам;
- оогамной - слияние неподвижной крупной яйцеклетки мелкими подвижными сперматозоидами.
У зигомицетов сливаются два, как правило, многоядерных гаметангия, морфологически хорошо отличимых от мицелия, на котором они образуются. Этот тип полового процесса получил название зигогамии.

Рис. 4. Типы полового процесса у грибов
Соматогамия - гаметы и половые органы отсутствуют, а сливаются обычные соматические клетки мицелия. У грибов с одноклеточными талломами часто наблюдается их слияние, называемое хологамией. Хологамия - одна из форм соматогамного полового процесса.
Среди грибов известны как гомоталличные, так и гетероталличные формы. У гомоталличных грибов к слиянию способны клетки одного мицелия. У гетероталличных сливаются только клетки разного полового знака, образующиеся на совместимых мицелиях.
Ядерные фазы в цикле развития грибов
У базидиомицетов и аскомицетов, половой процесс состоит из двух не совпадающих во времени этапов – плазмогамии и кариогамии.
У аксомицетов половые органы дифференцированы. Женский половой орган состоит из аскогона и нитевидной трихогины, через которую содержимое мужского полового органа - антеридия переливается в аскогон. При этом происходит только плазмогамия, ядра не сливаются, а образуют дикарион - ассоциацию двух ядер. Из оплодотворенного аскогона сразу же, без периода покоя, вырастают аскогенные гифы, в которых ядра дикариона синхронно делятся. На аскогенных гифах развиваются сумки. Ядра дикариона сливаются, диплоидное ядро делится мейотически, затем следует митотическое деление и вокруг восьми появившихся гаплоидных ядер формируются восемь аскоспор. Аскоспоры развиваются в сумках эндогенно.
У базидиомицетов после плазмогамии формируется дикариотический мицелий, существующий продолжительное время. На нем образуются базидии, в которых сначала сливаются ядра дикариона, а затем диплоидное ядро мейотически делится. На базидии экзогенно развиваются четыре базидиоспоры с гаплоидными ядрами.
Среди грибов известны как гомоталличные, так и гетероталличные формы. У гомоталличных грибов к слиянию способны клетки одного мицелия. У гетероталличных сливаются только клетки разного полового знака, образующиеся на совместимых мицелиях.
Характер смены ядерных фаз в цикле развития грибов может быть различен. У одних - гаплобионтов - редукционное деление диплоидного ядра происходит при прорастании зиготы. У других - диплобионтов - вся жизнь проходит в диплоидной фазе, а диплоидное ядро редукционно делится только перед образованием гамет. У аксомицетов и базидиомицетов в цикле развития имеется дикариотическая фаза, неизвестная у других групп.
Грибы из класса дейтеромицетов не имеют полового процесса, и весь их жизненный цикл происходит в гаплоидной фазе.
Принципы классификации и систематика грибов
В подразделении их на классы наиболее важными являются следующие признаки:
- количество, строение и расположение жгутиков;
- характер развития спор полового размножения;
- типы полового процесса и бесполого размножения;
- состав полисахаридов клеточных стенок.
Базируясь на современных представлениях о филогении грибов могут быть выдлены следующие классы:
Класс хитридиомицеты - Chytridiomycctes. Мицелий развит слабо или вегетативное тело представляет одиночную, иногда лишенную стенки клетку. Бесполое размножение при помощи зооспор с одним бичевидным задним жгутиком. Половой процесс - гаметогамия разных типов или хологамия. В клеточных стенках содержатся хитин и глюканы.
Класс гифохитриомицеты - Hyphochytriomycetcs. Вегетативное тело представляет собой одиночные клетки, иногда голые, у многих образуется ризомицелий. Бесполое размножение при помощи зооспор с одним перистым передним жгутиком. В клеточных стенках содержатся хитин и целлюлоза.
Класс оомицеты - Oomycetes. Мицелий хорошо развит, неклеточный. Бесполое размножение при помощи зооспор с двумя жгутиками - бичевидным и перистым. Половой процесс - оогамия. Клеточные стенки содержат целлюлозу и глюканы.
Класс зигомицеты - Zygomycetes. Мицелий хорошо развит, у большинства представителей класса он неклеточный. Бесполое размножение преимущественно спорангиоспорами. Половой процесс - зигогамия. Клеточные стенки мицелия содержат хитин и хитозан.
Класс аскомицеты - Ascomycetes. Мицелий хорошо развит, клеточный. Бесполое размножение при помощи конидий. Половой процесс - гаметангиогамия. Споры полового размножения образуются эндогенно - в сумке. Клеточные стенки содержат хитин и глюканы; у дрожжей - глюканы и маннаны (содержание хитина снижено).
Класс базидиомицеты - Basidiomycetes. Мицелий хорошо развит, клеточный. Бесполое размножение при помощи конидий. Половой процесс - соматогамия. Споры полового размножения образуются экзогенно - на базидии. Клеточные стенки содержат хитин и глюканы.
Класс дейтеромицеты, или несовершенные грибы - Deuteromycetes. Мицелий хорошо развит, клеточный. Бесполое размножение при помощи конидий. Половой процесс отсутствует. В клеточных стенках содержатся хитин и глюканы.
Кроме перечисленных классов среди грибов известны группы с неясным систематическим положением, например трихомицеты - Trichomycetes.
В современной микологии полифилетическое происхождение грибов часто находит отражение в подразделении их на ряд самостоятельных отделов соответственно основным эволюционным линиям. Это отделы Oomycota (Saprolegniomycofa), Chytridiomycota и Eumycota. Поскольку с настоящее время между отделами Chytridiomycota и Eumycota на основе сходства по ряду биохимических признаков предполагается филогенетическая связь, логично объединить их в один отдел Eumycota.
Классы грибов распределяются по этим отделам следующим образом.
Отдел Eumycota:
Классы: Chytridiomycetes, Zygomycetes, Ascomycetes, Basidiomycetes, Deuteromycetes.
Отдел Oomycota (Saproiegniomycota):
Классы: Oomycetes, Hyphochytriomycetes.