Лекция 8. Нервная ткань.
Общая характеристика и значение нервной
ткани.
Нервная ткань формирует нервную систему. Нервная система регулирует и координирует физиологические процессы
отдельных клеток, тканей, органов, их систем и организма в целом, хранит
информацию (память), перерабатывает и интегрирует следы памяти и сигналы из
внешней и внутренней среды.
Способность отвечать на раздражение – раздражимость - характерное свойство нервной ткани.
Нервная ткань, как и другие ткани, состоит из клеток и
межклеточного вещества. Клетки нервной ткани делятся на два вида: нейроны (нейроциты) и клетки глии (глиоциты). Нервная система человека содержит около 1012
нейронов, 1013 глиальных клеток и более 1013 синапсов.
Число клеточных типов неизвестно (не менее 100). Это множество образует сложную
пространственную структуру – единую сеть с многочисленными связями
как на уровне отдельной клетки, так и клеточных ансамблей (ЦНС).
У позвоночных животных и человека нервная
ткань составляет основу нервной системы, которая подразделяется на центральную и периферическую. ЦНС
включает головной и спинной мозг, содержащие большое
количество нервных клеток разного типа. ПНС включает нервы и концевые аппараты.
Небольшое количество нервных клеток скапливается в нервных узлах - ганглиях,
которые находятся в разных частях тела и входят в периферический отдел нервной
системы. Таким образом, у позвоночных животных и человека нервная система
представлена головным и спинным мозгом, нервами, нервными узлами (ганглиями),
нервными окончаниями.
Нервную систему делят на
соматическую, иннервирующую органы движения, и автономную, которая иннервирует
все внутренние органы. Обе части
тесно связаны в своей деятельности. Автономная нервная система по своим
анатомическим и физиологическим особенностям делится на симпатический и
парасимпатический отделы. Симпатические ядра находятся в боковых рогах спинного
мозга, начиная с I-го грудного до II-го поясничного сегмента. Ядра парасимпатического
отдела расположены в среднем, продолговатом мозге и крестцовом отделе спинного
мозга. Эфферентные нейроны парасимпатического отдела находятся во внутриорганных
ганглиях или нервных узлах, лежащих около иннервируемого органа. Эти системы отличаются по медиаторам, выделяемых в синапсах. Медиатором
парасимпатической нервной системы является ацетилхолин, медиаторы симпатической
– катехоламины (адреналин, норадреналин, дофамин).
Строение и классификация нейронов
Нейроны – главные клеточные элементы нервной ткани. У
млекопитающих и человека нейроны сосредоточены в сером веществе головного и
спинного мозга, а также в нервных узлах (ганглиях). Нейрон – возбудимая
клетка, которая воспринимает электрические сигналы, перерабатывает их и
передает различным органам тела, обеспечивает способность мозгв
к переработке и хранению воспринятой информации.
Нейрон состоит из тела и отростков. Тело (перикарион) и отростки аксон (нейрит), начинающийся от аксонного холмика, и дендриты – стандартные части нейрона.
Отростки, отходящие от перикариона, - аксон (нейрит)
и дендриты – участвуют в образовании синапсов. Существенная часть нейрона – цитоскелет.
Перикарион содержит цитоплазму, ядро, органоиды и специфические
структуры, присущие только ему. В центре перикариона
находится округлое ядро с малым количеством хроматина. Ядро содержит от одного
до трех ядрышек. Ядрышко крупное и резко базофильное.
 Нейрон содержит большое количество органоидов, что
связано с его высокой активностью. Хорошо развит комплекс Гольджи,
что свидетельствует о мощном транспорте белков, синтезируемых в гранулярной ЭПС. В цитоплазме нейронов много митохондрий,
что указывает на преобладание в них аэробного обмена. В некоторых нервных
клетках мозжечка есть центриоли, а в большинстве клеток головного и спинного
мозга они отсутствуют, что, очевидно, связано с их неспособностью к
митотическому делению. В цитоплазме находятся лизосомы и рибосомы, гранулы и
капли секрета, пигменты (липофусцин и меланин).
Нейрон содержит большое количество органоидов, что
связано с его высокой активностью. Хорошо развит комплекс Гольджи,
что свидетельствует о мощном транспорте белков, синтезируемых в гранулярной ЭПС. В цитоплазме нейронов много митохондрий,
что указывает на преобладание в них аэробного обмена. В некоторых нервных
клетках мозжечка есть центриоли, а в большинстве клеток головного и спинного
мозга они отсутствуют, что, очевидно, связано с их неспособностью к
митотическому делению. В цитоплазме находятся лизосомы и рибосомы, гранулы и
капли секрета, пигменты (липофусцин и меланин).
7Рис .35. Схема нейрона: 1 – тело клетки с
ядром; 2 – дендриты; 3 – аксон; 4 –оболочка; 5 – концевые разветвления;
6 – коллатераль; 7 – аксонный холмик
Аксонный холмик
– область перикариона, содержащая много микротрубочек
и нейрофиламентов, место, где начинается аксон и
генерируется потенциал действия.
Аксон (нейрит) (axis – ось) в клетке один и имеет
длину
от
Дендриты (dendron – дерево) – короткие, сильно ветвящиеся отростки.
Число их разное. Проксимальные области дендритов - продолжение перикариона, поэтому они содержат рибосомы, компоненты
гранулярной и гладкой ЭПС, элементы комплекса Гольджи.
Дедрты интегрально образуют до 95% всей рецепторной
поверхности (рецептивного поля) нейрона.
Они растут и подвергаются ретракции в
ответ на различные воздействия. Дендриты являются наиболее чувствительными
индикаторами функционального состояния нервных связей и любых изменений в
нервной системе. Дендриты в отличие от аксонов способны генерировать не только
потенциалы действия, но и градуальные генераторные
потенциалы.
Аксон и дендриты имеют сходное строение,
но различаются по функциям: аксон проводит импульс от перикариона,
а дендрит – к перикариону. Таким образом, возбуждение
по нейрону проходит от дендрита к перикариону и от него по аксону. В противоположном
направлении нервные импульсы не проходят, а тормозятся.
Цитоскелет нейронов состоит из микротрубочек, промежуточных филаментов (нейрофиламентов) и микрофиламентов. Микротрубочки – наиболее крупные элементы цитоскелета, их диаметр достигает 24 нм.
С ними связывают внутриклеточный, в т.ч. аксонный транспорт. Микротрубочки в перикарионе
и дендритах (в отличие от аксона) не имеют направленной ориентации. Большинство
микротрубочек аксона (+) концом направлено к терминали, а (-) концом – к перикариону.
Характер ориентации микротрубочек имеет важное значение для распределения по отросткам различных
органелл. К (+) полюсу перемещаются митохондрии и
секреторные пузырьки, а к (-) концу – рибосомы, мультивезикулярные тельца,
элементы комплекса Гольджи.
Классификация нейронов. Нейроны отличаются по размерам, форме перикариона, числу отростков, их синаптическим
связям, характеру ветвления дендритов, электрофизиологическим характеристикам,
химии нейромедиаторов, позиции в функциональных сетях
и т.д. В связи с этим классификации нейронов многочисленны
Морфологическая классификация нейрона определяется количеством отходящих от него отростков.
По этому признаку различают нейроны: мультиполярные –
с большим количеством отростков, биполярные – с двумя отростками, униполярные –
с одним отростком. Униполярные и биполярные нейроны округлые или овальные, а мультиполярные имеют неправильную многоугольную форму.
Мультиполярные нейроны
имеют число отростков больше двух. Среди этих отростков один аксон, остальные –
дендриты. Примером мультиполярного
нейрона могут служить мотонейроны передних рогов
спинного мозга.
Биполярные нейроны имеют аксон и
ветвящийся дендрит. По дедриту возбуждение проводится
из периферии в клетку, по аксону оно
направляется в мозг. Примером биполярных нейронов являются обонятельные
рецепторные нейроны.
Униполярные нейроны имеют один отросток, который распадается на две
ветви. Одна из них направляется к органу, а другая – в ЦНС. Такие нейроны
характерны для спинальных ганглиев высших позвоночных животных.
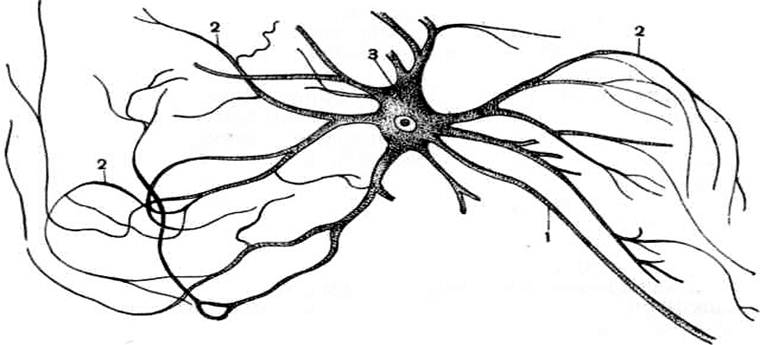
Рис.36. Мультиполярный нейрон
из спинного мозга зародыша коровы:
1 – аксон; 2 – дендриты; 3 – тело клетки с ядром.
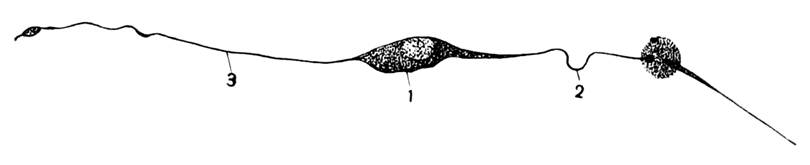
Рис.37. Биполярный нейрон из кожи
личинки стрекозы:
1 – тело клетки; 2 – дендрит; 3 – аксон.
Псевдоуниполярные нейроны
на самом деле обладают двумя отростками, которые на некотором расстоянии от перикариона соединяются между собой, образуя единый вырост.
 Рис. 38 . Униполярный нейрон:
Рис. 38 . Униполярный нейрон:
1 – тело клетки; 2 – ядро; 3 – отростки.
Аполяры – нейроны, у которых отсутствуют отростки. Условно к
ним относятся ранние нейробласты.
Позиция в нейронной цепочке позволяет
выделить 3 группы нейронов: чувствительные - передающие нервные импульсы к центральным отделам
нервной системы; двигательные – передающие возбуждение от центральных отделов
нервной системы к рабочим органам; вставочные (ассоциативные в нейронных сетях) – осуществляют связи между
нервными клетками.
Направление возбуждения также находит выражение в классификации нейронов: афферентные
нейроны – направление возбуждения к центру; эфферентные нейроны двигательных путей и трактов –
направление возбуждения от центра.
Химия нейромедиатора. Вещество, обеспечивающее передачу возбуждения,
называется медиатором. Обычно в каждом нейроне вырабатывается один основной нейромедиатор. При этом к названию нейромедиатора
добавляют окончание эргический.
По этой классификации различают нейроны: холинэргические – нейромедиатор ацетилхолин. К ним относятся двигательные
нейроны передних рогов спинного мозга, иннервирующие скелетные волокна;
парасимпатические нейроны блуждающего нерва, иннервирующие сердце, ГМК, железы
желудка;
Адренэргические - нейромедиатор –
норадреналин. К ним относятся постганглионарные
нейроны симпатического отдела вегетативной нервной системы, иннервирующие
сердце, ГМК сосудов и внутренних органов.
Длина аксона. Аксоны пирамидных нейронов коры больших полушарий достигают длины 50-
Модальность
– характер воспринимаемого и передаваемого сигнала (например, механорецепторные, зрительные, обонятельные нейроны и т.д.)
Отдел нервной системы. Выделяют нейроны вегетативного отдела нервной
системы и нейроны соматического отдела - чувствительные и двигательные, не
относящиеся к вегетативным.
Специфические
органоиды нервной клетки
 Специфическими образованиями нервной клетки являются тигроидное вещество и нейрофибриллы. Тигроидное
вещество (тигроид, вещество Ниссля) находится в перикарионе и
дендритах, его нет в аксоне и аксоном холмике. Под световым микроскопом тигроид выявляется как
скопление базофильного вещества в виде глыбок или зерен. Крупные глыбки
придают цитоплазме пятнистый вид шкуры тигра. С помощью электронного микроскопа
установлено, что тигроид
представляет собой мощно развитый гранулярный ЭПР. Ретикулум состоит из системы мембран с большим количеством
рибосом. Высокое содержание РНК обусловливает базофилию
Специфическими образованиями нервной клетки являются тигроидное вещество и нейрофибриллы. Тигроидное
вещество (тигроид, вещество Ниссля) находится в перикарионе и
дендритах, его нет в аксоне и аксоном холмике. Под световым микроскопом тигроид выявляется как
скопление базофильного вещества в виде глыбок или зерен. Крупные глыбки
придают цитоплазме пятнистый вид шкуры тигра. С помощью электронного микроскопа
установлено, что тигроид
представляет собой мощно развитый гранулярный ЭПР. Ретикулум состоит из системы мембран с большим количеством
рибосом. Высокое содержание РНК обусловливает базофилию
тигроида. В нем содержится и белок.
Рис. 39. Тигроидное вещество в мультиполярной
нервной
клетке спинного мозга.
Тигроид – обязательный компонент нервной клетки, легко
меняющийся в зависимости от функционального состояния. Тигролиз
– распыление тигроидного вещества, отражает глубокие
дистрофические изменения при нарушении целостности нейронов. При сильном
возбуждении нейрона тигроид может исчезнуть вообще.
Уменьшение тигроида и изменение его положения в
нейронах наблюдается также в результате патологических процессов: воспаления,
дегенерации, интоксикации. Все это дает основание рассматривать количество тигроида, форму его глыбок,
характер их расположения как показатели физиологического состояния нейрона.
В цитоплазме нейронов обнаруживаются нейрофибриллы
– нитчатые структуры. В теле нейрона и дендритах они образуют густую сеть.
В аксоне они вытягиваются по длине. Открытие нейрофибрилл привело к
возникновению нейрофибриллярной теории проведения нервного возбуждения.
Сторонники этой теории считали, что нейрофибриллы являются беспрерывным
проводящим элементом нервной системы, с чем связана ее главная функция. В
дальнейшем было установлено, что нейрофибриллы не принимают участие в процессе
проведения нервного возбуждения и прерываются в области контакта нервных
клеток. В настоящее время нейрофибриллы рассматривают артефактом, который
возникает посмертно в результате
осаждения серебра на структурах цитоскелета нейрона (нейрофиламенты, нейротубулин).
По современным представлениям, в
соответствии с нейронной теорией в проведении нервного возбуждения основная
роль принадлежит плазмалемме нейрона. Вопрос о значении нейрофибрилл остается
неясным. По слипанию нейрофибрилл определяют патологическое состояние нервной клетки.
Показано, что при старческом слабоумии наблюдается слипание и огрубление
нейрофибриллярной сети.. Часто в старческом возрасте и
при дистрофических процессах в нейронах обнаруживают зерна липофусцина. Их
рассматриваю остаточными тельцами, которые возникают в результате неполного
переваривания. Их накопление может привести к нарушению метаболизма в клетках и
к их гибели.
Нейросекреторные клетки
В определенных отделах мозга беспозвоночных и
позвоночных животных имеются нейроны, содержащие гранулы секрета. Такие
секретирующие нейроны называются нейросекреторными. Они имеют физиологические признаки
нейрона, но обладают выраженными признаками железистых клеток.
Нейросекрет синтезируются в связи с тигроидной субстанцией гранулярной ЭПС, оформляется в виде
секрета в системе аппарата Гольджи. Секрет
продвигается по аксону и выделяется из клеток в области их концевых
разветвлений. В отличие от обычных нейронов секрет высвобождается не в области
синапса, а в кровь или ликвор.
У низших позвоночных нейросекреторные клетки
расположены в преоптическом ядре головного мозга. У
высших позвоночных имеются два ядра в гипоталамической области. Аксоны
нейросекреторных клеток направляются в нейрогипофиз и
промежуточную долю аденогипофиза, образуя с ними
единую систему. Выделяемый нейросекреторными клетками продукт рассматривают как
гормон, регулирующий деятельность некоторых желез внутренней секреции и гонад,
где нервная регуляция оказывается редуцированной.
У беспозвоночных нейросекреторные гормоны
воздействуют на многие функции. У насекомых нейросекреция регулирует метаморфоз
(гормон окукливания, приводящий к лизису органов во время метаморфоза). У
червей, моллюсков, членистоногих нейрогормон имеет хроматофорную функцию, влияющую на окраску животного.
Активность нейросекреции связана во многих случаях с циклом жизни животного и
сезоном.
Нейроглия,
ее строение и классификация. Взаимоотношения
нейронов и нейроглии
Терминн «нейроглия» (glia –клей) ввел в обиход немецкий патологоанатом Рудольф
Вирхов для описания связывающих элементов между нейронами. Эти клетки
составляют половину объема мозга.
Нейроглия – вспомогательная и очень важная составная часть
нервной ткани, связанная с нейронами. По мере специализации нейрона как
индивидуальной клетки в процессе эволюции возникла организация более высокого
порядка – межклеточное «сообщество» нейрона и нейроглии. Нейроглия не принимает
непосредственного участия генерации и проведении нервных импульсов и, тем не
менее,
нормальное функционирование нейрона невозможно в
отсутствии или при повреждении глии. Нейроглиальные
клетки выполняют опорную, изолирующую, разграничительную, трофическую,
гомеостатическую, защитную функции. Клетки нейроглии не образуют синапсов.
По морфологическим и физиологическом свойствам и
происхождению нейроглию делят на макро- и микроглию.
Клетки макроглии подразделяются на эпендимоциты, астроциты, олигодендроциты, которые соответственно образуют эпендиму, астроглию и олигодендроглию.
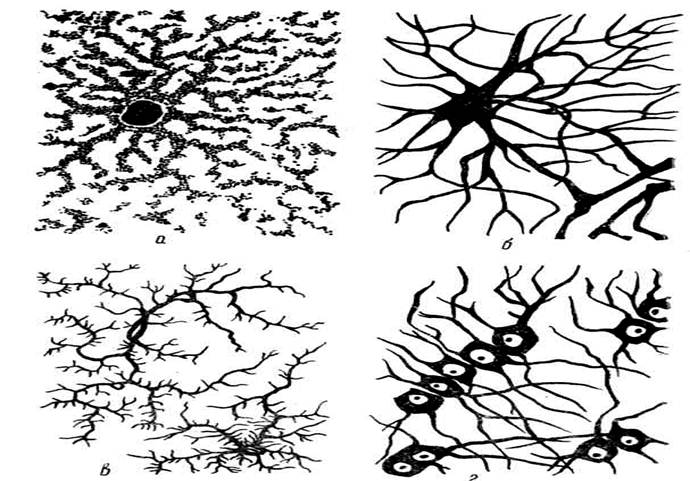
Рис.40. Различные
виды нейроглии:
а – плазматические
астроциты; б –
волокнистые астроциты;
в – олиго-дендроглиоциты; г – глиальные макрофаги.
 Наиболее древним видом макроглии
является эпендима. Клетки эпендимной глии эпендимоциты представляют
собой покрытые ресничками кубической формы, образующие выстилку желудочков
мозга и центрального канала головного и спинного мозга. Реснички эпендимных клеток обращены в полость мозга и их мерцанием
создается ток цереброспинальной жидкости. С возрастом количество ресничек на
свободной ничек на поверхности эпендимоцитов
уменьшается.временем во уменьшается.
Наиболее древним видом макроглии
является эпендима. Клетки эпендимной глии эпендимоциты представляют
собой покрытые ресничками кубической формы, образующие выстилку желудочков
мозга и центрального канала головного и спинного мозга. Реснички эпендимных клеток обращены в полость мозга и их мерцанием
создается ток цереброспинальной жидкости. С возрастом количество ресничек на
свободной ничек на поверхности эпендимоцитов
уменьшается.временем во уменьшается.
Рис .41.
Эпендима желудочка мозга:
1 – реснички, 2 – эпендимоцит
Ядра эпендимоцитов
овальные, богатые хроматином. В цитоплазме много митохондрий, хорошо развиты
комплекс Гольджи и гранулярная
ЭПС. Некоторым клеткам эпендимы характерна секреторная деятельность.
Предполагают, что выделяемый ими секрет входит в состав цереброспинальной
жидкости.
Астроглия выполняет опорную и трофическую функции. Астроглию образуют звездчатые клетки – астроциты
с длинными отростками. Астроциты оплетают нейроны, их
отростки, синапсы, кровеносные капилляры, изолирую и
структурно связывая их. Маркер астроцитов -
глиальный фибриллярный кислый белок промежуточных филаментов.
Астроциты имеют β-адренорецепторы и рецепторы многих нейромедиаторов.
Астроциты делятся на плазматические и волокнистые.
Плазматические астроциты расположены в сером веществе
мозга, имеют многочисленные короткие ветвящиеся отростки. Волокнистые астроциты находятся в белом веществе мозга, они имеют слабо - или неветвящиеся отростки.
Оба вида астроцитов заполняют все пространства между
нейронами и образуют пограничные мембраны между мозгом и мягкой мозговой
оболочкой.
Астроциты располагаются в мозгу таким образом, что со всех
сторон окружают и изолируют от посторонних афферентных влияний рецепторные
поверхности нейронов и нервные окончания. В этом заключается изолирующая
функция глии. Благодаря такой изоляции от
контакта с неспецифическими аксонными окончаниями
обеспечивается дискретный, локальный характер распределения импульсов.
При повреждении вещества мозга клетки
нейроглии и в особенности астроциты начинают
делиться, пролиферировать и замещать погибшие клетки, формируя рубец. Это – репаративная функция нейроглии. Установлено,
что астроциты
выполняют и другие многочисленные функции:
а) в гистогенезе они образуют проводящие пути для
миграции недифференцированных нейронов в коре мозжечка и для врастания аксонов
в зрительный нерв;
б) транспорт метаболитов из капилляров мозга в нервную
ткань;
в) регулируют химический состав межклеточной жидкости;
участвуют в метаболизме глутамата и ГАМК,
соответственно возбуждающего и тормозного нейромедиаторов ЦНС. После высвобождения этих нейромедиаторов в синаптическую
щель часть молекул поступает в астроциты, где они
превращаются в глутамин;
г) изолируют рецептивные поверхности нейронов;
д) принимают участие в фагоцитозе и патологических
процессах (пролиферация и замещение погибших нейронов);
е) выделяют вещества, способствующие росту аксонов –
фактор роста нервов, компоненты межклеточного матрикса ламилин
и фибронектин, ингибирующие и ускоряющие рост
отростков нейронов;
ж) предшественники астроцитов
контролируют направление миграции нейробластов и
формирование и отростков;
з) стабилизируют структуры ЦНС, ингибируя рост
отростков неронов и предотвращая их апоптоз;
и) обеспечивают ионный обмен, особенно ионов К.+
Олигоденроглия состоит из клеток олигодендроцитов.
В отличие от астроцитов они характеризуются наличием
небольшого количества коротких отростков. Форма клетки удлиненная, ядро
округлое с одним ядрышком. Олигодендроциты находятся
в сером и в белом веществе головного и спинного мозга, а также за пределами
ЦНС. Этих клеток много в коре головного мозга, зрительном бугре, продолговатом
мозге, спинальных ганглиях. В сером веществе мозга олигодендроциты
находятся в непосредственном контакте с перикарионами и отростками нейронов. В белом веществе они
расположены рядами между нервными волокнами. Олигодендроциты
ЦНС относятся к миелинобразущим клеткам и участвуют в
формировании нервных волокон.
К олигодендроцитам относятся
глиоциты-сателлиты, которые расположены на
поверхности нейронов и леммоциты, или шванновские клетки, образующие оболочки нервных
волокон. Маркер шванновских клеток – белок S 100 В.
Олигодендроциты синтезируют липоидное вещество миелин, который входит в
состав плазмалеммы. Миелину присущи изоляционные свойства. Изолируя отростки нервных
клеток, олигодендроциты препятствуют рассеиванию
нервного импульса. Они формируют оболочки вокруг нейронов, обеспечивая
барьерно-транспортные функции. Они принимают участие в питании нейронов и
водном обмене мозга. Вместе с астроцитами они
участвуют в хранении следов возбуждения в нервной системе.
Микроглия состоит из клеток, называемых микроглиоцитами,
которые рассеяны по нервной ткани. Особенно часто они встречаются около
сосудов. Клетки микроглии имеют округлую форму с коротким ветвящимися отростками. Они активно передвигаются
по нервной ткани и проявляют фагоцитарную активность, поглощая как гибнущие
нейроны и нервные окончания, так и бактерии. При движении эти клетки могут
менять форму. Микроглию рассматривают как защитный
элемент нервной ткани. Микроглиоциты осуществляют
активацию иммунологических ответов и стимуляцию регенерации нейронов и их
отростков.
Мантийные глиоциты располагаются в периферических нервных узлах вокруг перикарионов. Они образуют вокруг них глиальные капсулы.
Клетки имеют уплощенную форму с маленьким ядром и хорошо развитым ядрышком.
Хорошо развит цитоскелет. Мантийные глиоциты выполняют изолирующую, трофическую, опорную,
защитную и барьерную функции.
Нервные волокна
Тела нейронов образуют серое вещество головного и
спинного мозга, а также нервные ганглии беспозвоночных и позвоночных животных.
Связь ЦНС и ганглиев с органами осуществляется при помощи проводящих элементов
– нервов, основу которых составляют нервные волокна.
Отростки нервных клеток, окруженные плазмолеммой олигодендроцитов или шванновских
клеток, называются
нервными волокнами. Отросток нервной клетки в составе нервного
волокна называется осевым цилиндром, а глиальные клетки, формирующие оболочку
волокна – леммоцитами, или
шванновскими клетками.
Нервные волокна образуют в головном и спинном мозге
проводящие пути, а на периферии – нервы. В пределах ЦНС нервные волокна входят
в состав белого вещества мозга. По нервным волокнам осуществляется проведение
нервных импульсов.
Толщина соматических нервных волокон равна 12-14 мкм,
автономных - 5-7мкм. В зависимости от строения покрывающих оболочек нервные
волокна подразделяются на два вида: безмякотные (немиелиновые) и мякотные (миелиновые).
Безмякотные (немиелиновые) нервные
волокна входят в
состав периферических нервов, идущих к внутренним органам, но многие сенсорные
волокна также являются безмякотными. Они имеют несколько осевых цилиндров
(3-5, иногда до 12), окруженных шванновскими
клетками.
В электронных микрофотографиях видно, что
каждый осевой цилиндр погружен в леммоцит, ее
клеточная мембрана смыкается и образует мезаксон – сдвоенные мембраны шванновской
клетки. Каждая шванновская клетка подобным образом
окружает несколько осевых цилиндров. В зависимости от числа осевых цилиндров,
погруженных в леммоцит, может быть разное количество мезаксонов в нервном волокне. Миелин отсутствует. Шванновские клетки на всем протяжении окутывают безмякотное волокно, препятствуя его соприкосновению с
окружающей средой.
Поскольку отростки нервных клеток покрыты плазмалеммой
шванновских клеток только один раз, то нервный
импульс при прохождении рассеивается. Он проходит по безмякотным
нервным волокнам в 10 раз медленнее, по сравнению с мякотными.
По безмякотным волокнам импульс проходит со скоростью
меньше 1м/с.
 Рис.42.
Схема строения немиелинового нервного волокна:
Рис.42.
Схема строения немиелинового нервного волокна:
1 –
ядро; 2 – цитоплазма леммоцита; 3-4 – аксоны нервных
клеток; 5 – митохондрия; 6 - гранулярная эндоплазматическая сеть; 7 – микротрубочки; 8 –
коллагеновые волокна эндоневрия; 9 – базальная
мембрана; 10 – мезаксон
Мякотные нервные волокна составляют белое вещество головного и спинного мозга
и входят в периферические нервы. Мякотное нервное волокно состоит из одного
осевого цилиндра, вокруг которого шванновские клетки
образуют миелиновую оболочку. Нервное волокно, состоящее из одного осевого
цилиндра и расположенных вокруг него шванновских
клеток, называют мякотным, или миелиновым.
 Рис. 43.
Схема строения миелинового волокна:
Рис. 43.
Схема строения миелинового волокна:
1–аксон;
2–мезаксон; 3–насечки; 4–перехват; 5
-цитоплазма леммоцита; б–ядро леммоцита; 7–неврилемма; 8 –
эндоневрий.
Характерная
особенность шванновских клеток – наличие в них
липоидного вещества миелина, которое образует вокруг осевого цилиндра мякотную
(миелиновую) оболочку. Каждая шванновская клетка миелинизирует небольшой сегмент только одного аксона.
Мякотная, или миелиновая, оболочка примыкает к осевому цилиндру и окружает его
чехлом. Она выполняет роль изолятора. Этим объясняется
большая скорость проведения нервных импульсов мякотными нервными волокнами,
т.к. скорость проведения возбуждения зависит от диаметра и степени миелинизации нервного волокна.
Миелин регулярно прерывается через
определенные промежутки. Фактически эти участки, лишенные миелина, являются
границами между двумя соседними глиоцитами, где они
соединяются при помощи коротких отростков и называются узлами нервного волокна (перехваты Ранвье).
В перехвате Ранвье аксолемма осевого цилиндра не покрыта
миелиновой оболочкой. Аксолемма перехватов Ранвье содержит множество потенциалозависимых
Nа+-каналов,
необходимых для поддержания импульсной активности. Эти каналы практически
отсутствуют в покрытых миелином сегментах аксона.
Скачкообразное проведение нервных импульсов в
миелиновых волокнах, когда сигнал перескакивает от одного перехвата к
другому, обеспечивают Nа+- каналы перехватов Ранвье.
По этой же причине в миелиновых волокнах (в отличие от не имеющих перехватов немиелиновых волокон) скорость проведения нервных импульсов
выше.
Участок между узлами называется межузловым сегментом. В периферических нервах
каждый миелиновый сегмент пересекается с воронкообразными щелями, идущими в
косом направлении от наружной поверхности оболочки к
внутренней. Они называются «насечками миелина» (шмидтлантермановскими насечками). Шмидтлантермановские
насечки – это участки расслоения миелина, образовавшиеся при миелинизации. Функция насечек неясна. В зависимости от
длины миелинового сегмента количество насечек миелина бывает различным. Они
отсутствуют в пределах ЦНС. Существует мнение, что насечки являются
артефактами, возникающими посмертно в ходе фиксации препаратов.
Осевой цилиндр содержит митохондрии,
элементы гладкой ЭПС и цитоскелета – микротрубочки, нейрофиламенты и микрофиламенты.
Скорость проведения нервного импульса зависит от диаметра аксона, а сам диаметр
определяется количеством содержащихся в нем нейрофиламентов. В нормальных и патологических условиях
количество нейрофиламентов и диаметр аксона тесно коррелируют.
Нервные волокна имеют микротрубочки, по
которым перемещаются вещества в клетку и из клетки. Этот процесс называется аксотоком или аксонным
транспортом. Различают быстрый (100-1000 мм/сутки) и медленный (1-10 мм/сутки) аксонный транспорт, а также антероградный
(транспорт от перикариона) и ретроградный (к перикариону). Основной материал антероградного
транспорта - белки, синтезированные в перикарионе
(например, белки ионных каналов, ферменты синтеза нейромедиаторов).
За счет ретроградного транспорта в клетку поступают различные нейротропные вещества, оказывающие трофическое влияние на
нейрон. Направление аксотока обеспечивается за счет
полярности микротрубочек и осуществляется кинезинами
и динеинами.
Внешняя плазмалемма шванновских
клеток окружена базальной мембраной.
Наличие в мякотном нервном волокне только
одного отростка нервной клетки, изолированного миелиновой оболочкой, а также
узловых перехватов, обеспечивает быстрое (до 120 м/с) и точное проведение
нервных импульсов.
Выше изложено строение мякотного
периферического нервного волокна. Мякотные нервные волокна ЦНС построены
сходным образом, однако их оболочка образована не леммоцитами,
а олигодендроцитами. Насечки и перехваты в них отстствуют, нет и базальных мембран.
Нервные стволы
Нервные стволы (нервы) образованы пучками мякотных и безмякотных
нервных волокон, которые объединяются соединительной тканью, образующей
соединительнотканные оболочки. В нерве может быть множество волокон только
мякотных или только безмякотных. Есть нервы, в
которых встречаются и те и другие. Наружная оболочка нерва – эпиневрий - состоит из волокнистой
соединительной ткани, объединяющей все пучки в составе нерва. Периневрий –
соединительнотканная оболочка, окружающая каждый отдельный пучок нервных
волокон. Эндоневрий – рыхлая
соединительная ткань между отдельными нервными волокнами. Эта ткань
связывает нервные волокна в пучки,
соединяясь с их базальной мембраной.
Периневриальный барьер необходим для поддержания гомеостаза в эндоневрии. Барьер контролирует транспорт молекул через периневрий к нервным волокнам, предотвращает доступ в эндоневрий инфекционных агентов.
Периферический нерв содержит разветвленную
сеть кровеносных сосудов. В эпиневрии и в наружной
части периневрия содержатся артериолы и венулы, а также лимфатические сосуды. В эндоневрии
проходят кровеносные капилляры. Периферический нерв иннервирован – имеет
специальные нервные волокна.
Соединение нейронов между собой (синапсы)
Нейроны, как отдельные единицы нервной системы,
функционируют не изолированно. Они соединены между собой и образуют единую
сеть, которая передает возбуждение от рецепторов в ЦНС и от нее в различные
органы.
Специфические контакты нейронов между собой, а также нейронов с клетками испольнительных органов, называются синапсами. Синапсы обеспечивают передачу возбуждения от одной нервной клетки другой, а также от нейронов к рецепторам или эффекторам. Несмотря на разнообразие синапсов, в их строении имеются общие черты. В синапсе выделяют пресинаптическую и постсинаптическую мембраны и пространство между ними – синаптическую щель шириной 20-30 нм.
Рис. 44. Схема ультрамикроскопического строения различных типов синапсов:
А – цитотопография синапсов; Б – синапс
тормозного типа; В – синапс возбудительного типа; Г – электротонический
синапс; 1 – асосоматический
синапс; 2 – аксодендритические синапсы;
3 – аксоаксональный синапс; 4 – дендриты; % -
дендритный шипик; 6 – аксон; 7 – синаптические
пузырьки; 8 – пресинаптическая
мембрана; 9 – постсинаптическая
мембрана; 10 – синаптическая щель; 11 -
постсинаптические структуры.
Конечные участки аксонов и дендритов в области синапса
не имеют мякотной оболочки и расширены в пресинаптический
мешочек. Мешочек имеют синаптические пузырьки
диаметром 40-59 нм. Они содержат и секретируют медиаторы.В зависимости от
типа выделяемого медиатора различают синапсы:
а) холинэргические
– выделяют ацетилхолин;
б) адренэргические
– выделяют норадреналин, дофамин (катехоламины);
в) серотонинэргические –
выделяют серотонин;
г) пептидэргические –
выделяют пептиды (эндорфины, энкефалины)
и аминокислоты (глицин, глутамат, ГАМК).
В таких синапсах передача нервного импульса
осуществляется при помощи химического вещества – медиатора. Такие синапсы называются синапсами с химической передачей.
При изменении мембранного потенциала в терминалях нейромедиаторы выходят
в синаптическую щель через поры диаметром 4-5 нм, имеющиеся в пресинаптической мембране и связываются со своими
рецепторами в постсинаптической мембране, вызывая изменение мембранного
потенциала постсинаптического нейрона.
В пресинаптической части расположены синаптические
пузырьки и митохондрии. Пресинаптическая мембрана
содержит потензиалозависимые Са2+-каналы.
При деполяризации мембраны каналы открываются и ионы Са2+
входят в терминаль, запуская в активных зонах экзоцитоз нейромедиатора. Синаптические пузырьки содержат нейромедиатор.
Постсинаптическая мембрана содержит рецепторы нейромедиатора
и ионные каналы.
Синаптическая передача – сложный каскад событий. Она возможна при
реализации ряда последовательных процессов: синтез нейромедиатора,
его накопление и хранение в синаптических пузырьках
вблизи пресинаптической мембраны, высвобождение нейромедиатора из нервной терминали, кратковременное взаимодействие нейромедиатора с рецептором, встроенным в постсинаптическую
мембрану, разрушение нейромедиатора или захват его
нервной терминалью. Многие неврологические и
психические заболевания сопровождаются нарушениями синаптической
передачи.
В некоторых синапсах синаптическая
щель отсутствует и его структурной основой является плотный контакт. В таком
синапсе возбуждение может передаваться без участия медиатора, т.к. мембраны
клеток соприкасаются. Эти синапсы называются синапсами с электрической
передачей. В синапсах такого строения пресинаптическая
мембрана также имеет поры, но их в 5 раз меньше, чем в синапсах с химической
передачей возбуждения. Поры электрических синапсов являются межклеточными
диффузионными каналами, соединяющими соприкасающиеся клетки.
По структуре и локализации синапсы
подразделяются на 3 группы: межнейронные, рецепторно-нейрональные
и нейроэффекторные. Межнейронные синапсы
подразделяются на аксодендритические, аксосоматические
и аксоаксональные.
Межнейронные синапсы являются синапсами между двумя нейронами. Если аксон
одного нейрона контактирует с дендритом другого постсинаптического нейрона, то
такие синапсы называются аксодендритическими. Если аксон одного нейрона контактирует
с перикарионом другого постсинаптического нейрона, то
такой синапс называется аксосоматическим. Если же аксон
одного нейрона контактирует с аксоном другого постсинаптического нейрона, то
такой синапс называется аксо-аксональным.
Межнейронные синапсы очень многочисленны. На
поверхности перикариона и отростков одного
пирамидного нейрона в коре больших полушарий головного мозга имеется около 104
синапсов.
Рецепторно-нейрональные (рецепторно-дендритные)
синапсы являются синапсами между рецепторными клетками, сходными с нейронами,
специализированными эпителиальными, нейроглиальными клетками, с одной стороны,
и дендритами чувствительных нейронов – с другой. Примером синапсов такого типа
у позвоночных являются синапсы вкусовых
сосочков, боковой линии рыб, внутреннего уха, кожи, соединительной ткани.
Нейроэффекторные (аксоэффекторные) синапсы
являются синапсами между аксоном двигательных (эффекторных)
нейронов и клетками, не принадлежащими к нервной системе.
У человека и млекопитающих хорошо изучены
двигательные и секреторные нейроэффекторные синапсы,
или эффекторные нервные окончания. Первые
представляют собой синаптические соединения между
аксоном двигательного нейрона и поперечнополосатыми мышечными волокнами,
поперечнополосатыми и гладкомышечными клетками, а вторые – между аксонами
двигательного нейрона с секреторными клетками. Существуют синапсы между аксоном
эфферентного нейрона и другими клетками – жировыми, ресничными и др.
Концевые нервные аппараты и их
классификация
В ПНС нейроны образуют концевые нервные аппараты
(нервные окончания), которые интегрируют функции органов их систем,
обеспечивают взаимодействие организма с
внешней средой. Окончания периферических
нервов подразделяют на чувствительные (афферентные) двигательные
(эфферентные).
Приспособления, которые воспринимают
раздражения, называются рецепторными аппаратами, или
чувствительными нервными окончаниями, а нервы, проводящие возбуждение – чувствительными.
Реализация нервных импульсов осуществляется эффекторными аппаратами (двигательными нервным
окончаниями), а проведение возбуждения к ним происходит по двигательным нервам.
Концевые нервные аппараты – сложные
образования. В их состав входят не только нервные волокна, но и ткани, в
которых они оканчиваются. Структура концевых аппаратов разнообразна, меняется в
зависимости от условий, в которой они находятся.
 Эффекторный аппарат
хорошо представлен на примере двигательной бляшки. Он располагается на
поперечнополосатом мышечном волокне в виде разветвления осевого цилиндра
мякотного нервного волокна, которое теряет миелин. По данным электронной
микроскопии, для двигательной бляшки характерно отчетливое разграничение
нервной и мышечной частей. В гладких мышцах двигательная иннервация
осуществляется безмякотными нервными окончаниями.
Эффекторный аппарат
хорошо представлен на примере двигательной бляшки. Он располагается на
поперечнополосатом мышечном волокне в виде разветвления осевого цилиндра
мякотного нервного волокна, которое теряет миелин. По данным электронной
микроскопии, для двигательной бляшки характерно отчетливое разграничение
нервной и мышечной частей. В гладких мышцах двигательная иннервация
осуществляется безмякотными нервными окончаниями.
Рис.45.
Схема субмикроскопического строения
двигательного нервного
окончания на миоците:
1 – постсинаптическая мембрана миоцита; 2 – митохондрия; 5 – светлые пресинаптические пузырьки; 4 – пресинаптическая
мембрана нейрита
Секреторные окончания эффекторных нейронов
представлены аксонами, выступающими в синаптический
контакт с железистыми клетками. Концевые разветвления аксона либо подходят
вплотную к секреторной клетке, либо глубоко вдавливаются в нее. Нейролемма аксона и плазмалемма секреторной клетки образуют
соответственно пресинаптическую и постсинаптическую
мембраны, разделенные узкой синаптической щелью.
Холинорецепторы присутствуют также в мембране мышечного волокна вне
синапса, но здесь их концентрация на порядок меньше, чем в постсинаптической
мембране и обозначаются они как холинорецепторы.
Рецепторные аппараты (рецепторные нервные окончания). Рецепторные
(воспринимающие) нервные окончания у
позвоночных представляют собой концевые
аппараты дендритов чувствительных нейронов, тела которых располагаются чаще
всего в спинальных ганглиях и их
аналогах – черепномозговых чувствительных узлах или в
периферических вегетативных ганглиях. В зависимости от того, откуда они
воспринимают раздражение, различают экстерорецепторы, интерорецепторы
и проприорецепторы. Первые
воспринимают раздражения из внешней среды, вторые – с внутренних органов,
третьи – осуществляют мышечно-суставные чувства. Они
Экстерорецепторы расположены в коже,
слизистых оболочках ротовой полости, прямой кишки, наружных половых органов,
склере, роговице. Интерорецепторы локализованы во внутренних органах. С учетом
специфики раздражителя различают тактильные, холодовые,
тепловые, болевые рецепторы, барорецепторы,
хеморецепторы, механорецепторы.
По морфологическим особенностям рецепторные окончания
могут быть свободными, располагающимися между клетками иннервируемой
ткани, и несвободными, инкапсулированными (заключенными в
особые соединительнотканные капсулы).
Свободные нервные окончания – наиболее распространенный тип сенсорных рецепторов.
Большинство свободных нервных окончаний – механорецепторы.
Распространены в прослойках соединительной ткани внутренних органов, а также в
соединительнотканной основе кожи. Свободные нервные окончания эпидермиса
расположены в базальном и шиповатом слоях. В области кожи с высокой тактильной
чувствительностью (пальцы рук) терминали достигают
зернистого слоя. Некоторые окончания в эпидермисе специализированы для
регистрации изменений температуры. Свободные нервные окончания имеются и в
других органах чувств (слуха, равновесия, вкуса), закладывающихся из эктодермы.
В многослойном эпителии локализованы чувствительные
осязательные клетки Меркеля, имеющие округлую или
удлиненную форму. Они соединены с эпителиоцитами при
помощи десмосом и формируют контакт с нервными терминалями. В клетках Меркеля
обнаружены пептиды и нейроспецифические вещества, что
свидетельствует об их эндокринной функции. Это позволяет рассматривать их как
компонент диффузной нейроэндокринной системы.
Капсулированные чувствительные нервные окончания построены по единому плану и наблюдаются в
соединительной и мышечной тканях. Эти рецепторные нервные окончания покрыты
соединительнотканной капсулой (наружная капсула), а изнутри - нейроглией
(внутренняя капсула, или колба) Наружная капсула
образована плотной соединительной тканью.
К
капсулированным рецепторам мышечной
ткани относятся нервно-мышечные веретена и капсулированные
кустики. Они являются специфическими рецепторами соматической
мускулатуры, воспринимающие ощущение растяжения мышечного волокна.
 Рис.46. Капсулированные нервные окончания:
Рис.46. Капсулированные нервные окончания:
А -
продольный разрез пластинчатого тельца из кожи человека; Б - поперечный
срез пластинчатого тельца из кожи пальца
человека;
1 – нервное волокно; 2 - внутренняя луковица; 3 -
клетки пластинок капсулы
Одним концом они прикреплены к перимизию
мышечного волокна, а другим - к сухожилию. В гладкой мускулатуре внутренних
органов находятся кустиковидные свободные рецепторные
окончания.
 Рис.47. Осязательные тельца:
Рис.47. Осязательные тельца:
А – из сосочка
кожи пальца человека;
Б – из кожи пальца
человека;
1 – эпидермис; 2 – основное волокно; 3 – добавочное волокно; 4 –
расширение основного волокна; 5 – олигодендроглиоциты
(осязательные клетки); 6 – веточка нервного волокна, выходящая из
тельца; 7 – пластинка
Строение инкапсулированных рецепторных
окончаний изучены
на примере осязательных телец (телец Мейснера) и
пластинчатых
телец (телец Фатер - Пачини).
Осязательные
тельца расположены в сосочковом слое кожи и являются механорецепторами. Они воспринимают поверхностную
тактильную чувствительность.
Тельце
имеет удлиненную форму. Внутренняя часть тельца состоит из уплощенных
нейроглиальных клеток, окружающих дендрит и образующих вместе внутреннюю колбу
тельца. С внешней стороны тельце покрыто соединительнотканной капсулой и
образует наружную колбу. В теле человека наиболее распространены пластинчатые
тельца, или тельца Фатер – Пачини, которые являются механорецепторами
и воспринимают глубокую тактильную чувствительность. Они встречаются в
глубоких слоях кожи, на брыжейке, в
молочной железе, кишечнике, поджелудочной железе, соединительной ткани
внутренних органов, около кровеносных сосудов. Тельце имеет овальную форму и размеры в пределах 0,5 – 1,0 мм.Внутренняя колба, наружная
капсула ми терминальное нервное волокно – основные компоненты тельца.
Внутренняя колба тельца содержит нейроглиальные клетки. Вокруг внутренней колбы
находится мощная соединительнотканная капсула, состоящая из плоских серповидных соединительнотканных
клеток.
 Рис.48. Фатер-пачиниево тельце:
Рис.48. Фатер-пачиниево тельце:
Электронная микрофотограмма: 1 – аксон;
2 – митохондрии, 3 – пузырьки.
К тельцу Фат ер-Пачини
подходит толстое миелинизированное нервное волокно. Внутри наружной капсулы они образуют
несколько перехватов Ранвье. Подойдя к внутренней
колбе рецептора, нервное волокно теряет миелин и переходит в чувствительную
нервную терминаль. Эти тельца воспринимают ощущение
давления на органы и внутриорганное давление.
К механорецепторам примерно
такого же строения относятся луковицеобразные тельца (тельца Гольджи-Маццони), которые расположены в концевой части
сухожилий на границе с мышцей, а также в связках капсулы суставов и
воспринимают растяжение.
В теле человека встречаются концевые
колбы (колбы Краузе), которые являются терморецепторами. Они расположены в
соединительнотканной основе кожи, слизистых и серозных оболочках. Они также
имеют тонкую соединительнотканную капсулу, образующую наружную колбу рецептора.
Температурные раздражения воспринимают капсулированные клубочки (тельца Руффини) – крупные рецепторы веретеновидной формы длиной до
К группе капсулированных нервных окончаний относятся
генитальные тельца (тельца Догеля). Они обнаружены в
соединительной ткани половых органов, головки полового члена, клитора и других
частях тела. По своему строению они напоминают тельца колбы Краузе.
Генитальное тельце является механо- и барорецептором, поскольку
реагирует на изменение кровяного давления.
Из капсулированных механорецепторов
кожи птиц наиболее распространены тельца Хербста и тельца Грандри,
расположенные в восковице пластинчатоклювых. Тельце Хербста имеет такое же строение, как и тельца Фатер – Пачини. Тельце Грандри мельче телец Хербста и
они обладают более тонкой соединительнотканной капсулой. Внутри капсулы
находятся две крупные нейроглиальные клетки с крупными овальными ядрами.
Таким образом, инкапсулированные рецепторные окончания
всегда состоят из разветвлений осевого цилиндра чувствительного нейрона,
оканчивающихся на глиальных клетках, окруженных соединительнотканной капсулой.
Рефлекторная дуга и динамическая
поляризация нейронов
Все тканевые элементы нервной системы образуют
нейронные связи, благодаря которым осуществляется рефлекс - ответная
реакция организма на различные раздражения, осуществляемая при помощи нервной
системы. Рефлекс осуществляется с помощью рефлекторной дуги. Рефлекторная дуга
имеет следующие элементы: рецептор, чувствительный нерв, участок ЦНС,
двигательный нерв, исполнительный орган. При помощи рефлексов происходит
приспособление организма к меняющимся условиям окружающей среды.
Различают простые и сложные рефлексы. Рассмотрим
простейший рефлекс, осуществляемый в спинном мозгу без участия головного мозга.
Такой рефлекс осуществляется при учатии трех типов
нейронов: чувствительных, вставочных и двигательных.
Чувствительные нейроны- клетки, воспринимающие раздражение. У человека и
высших позвоночных животных они находятся в спинальных ганглиях, или узлах,
расположены по обеим сторонам спинного мозга, по ходу его задних корешков.
Здесь расположены чувствительные униполярные нейроны. От нейрона отходит один
отросток, который разветвляется на две ветви. Один из этих ветвей более
длинный, направляется по спинномозговому нерву на периферию, где он
заканчивается чувствительным концевым аппаратом, воспринимающим раздражение.
Другой более короткий отросток входит в спинной мозг и служит проводником
возбуждения от чувствительного концевого аппарата. В белом веществе этот
центральный отросток разветвляется.
Одна ветвь направляется вверх, а другая
– вниз. Пройдя некоторое расстояние, обе ветви входят в серое вещество и заканчивается
на телах нейронов, называемых вставочными (связывающими, промежуточными).
Вставочные нейроны – небольшие мультиполярные
клетки с короткими дендритами. Их единственный нейрит проникает в белое
вещество, где разделяется на две ветви, одна из которых направляется вверх, а
другая – вниз. В выше- и нижележащих отделах спинного мозга они опять заходят в
серое вещество и вступают в контакт с двигательными,
или моторными, нейронами. Этот тип связывающих нейронов характеризуется тем,
что их отростки не выходят за пределы спинного мозга и
объединяет только его отделы. Кроме таких клеток, в задних рогах и в средней
части серого вещества имеется и другой тип связывающих нейронов. Их восходящий
отросток отличается значительной длиной и поэтому достигает стволовой части
головного мозга.
Связующие клетки представляют второй тип
нейронов, принимающих участие в осуществлении рефлекса. В них происходит
трансформация чувствительного импульса в двигательный.
Дальнейший путь этого импульса связан с проводящими волокнами связующих
нейронов и наличием в спинном мозге двигательных нейронов. На теле этих нервных
клеток оканчиваются отростки вставочных нейронов.
Двигательные (моторные) нейроны расположены в передних рогах серого вещества спинного
мозга отдельными группами, которые называются ядрами. Эти – самые крупные
клетки спинного мозга. Они являются мультиполярными и
отличаются сильно разветвленными дендритами. Аксон этих клеток выходит из
спинного мозга по переднему корешку и направляется к мышце. Двигательный
импульс по волокнам этих клеток попадает к исполнительному органу, который
совершает работу. На этом заканчивается путь чувствительного импульса, который
возник в рецепторе.
Белое вещество состоит из волокон, большая
часть которых принадлежит к мякотным. Они расположены
вдоль спинного мозга и образуют проводящие пути – короткие,
объединяющие разные уровни спинного мозга, и длинные, соединяющие спинной мозг
с головным. Кроме описанной простой рефлекторной дуги, в организме высших
позвоночных (птицы и млекопитающих) преобладают сложные рефлекторные дуги. В
них через синапс чувствительного нейрона информация достигает от
нескольких до сотен и тысяч
ассоциативных нейронов. Они анализируют информацию и передают возбуждение к эффекторному нейрону. Сложные рефлекторные дуги
осуществляют переключение через
нейронные сети ядерных центров ствола, подкорковые ядра и экранные центры коры
головного мозга.
Гистогенез и регенерация элементов
нервной системы
Нервная ткань (за исключением микроглии) развивается из эктодермы. В дорзальной части
эктодермы в процессе эмбриогенеза формируется нервная пластинка. По
краям нервной пластинки возникают утолщения, называемые нервными валиками.
Нервная пластинка постепенно погружается вглубь зародыша, а нервные валики
перемещаются навстречу друг к другу. В результате соединения нервных валиков
нервная пластинка превращается в нервную трубку. Из нее в дальнейшем
развивается головной и спинной мозг.
В процессе нейруляции, в ходе которой образуется нейроэктодерма, формируются нервная трубка, нервный гребень
и нейрогенные плакоды. В нейроонтогенезе
происходит ряд морфогенетических процессов (гибель нейронов, направленный рост
аксонов и т.д.), совокупный эффект которых приводит к формированию нервной
системы.
Нервная трубка содержит внутреннюю пограничную мембрану, эпендимный
слой, плащевой (мантийный) слой, краевую вуаль, наружную пограничную мембрану.
Матричные клетки эпендимного слоя – источник почти
всех клеток ЦНС. Они сосредоточены вблизи внутренней пограничной мембраны.
Закончившие пролиферации клетки (нейробласты),
а также потенциально способные к пролиферации глиобласты
вытесняются в плащевой слой. Часть матричных клеток остается in situ -
будущая эпендима. Часть потомков выселяется из эпендимного
слоя в мантийный – будущее серое вещество.
Нейробласты дают начало всем нейронам ЦНС. Нейроны – классический
пример клеток, относящихся к статической популяции. Они in vivo не способны к пролиферации и
обновлению. Обонятельные нейроны выстилки носовых ходов – единственное
известное исключение.
Глиобласты – предшественники макроглии
(астроциты и олигодендроциты).
Все типы макроглии способны к пролиферации. Слой
клеток эпендимы до поздних стадий эмбриогенеза сохраняет способность к
образованию нейробластов и спонгиобластов.
Затем начинаются морфогенетические процессы –
индукция, адресная миграция клеток, направленный рост аксонов и гибель клеток,
нейротрофические взаимодействия – хорошо выражены в развивающейся нервной
системе. Жесткость организации мозга определяют два момента: адресная миграция
клеток и направленный рост их отростков.
Адресная миграция клеток – обязательное условие формирования мозга. Наиболее
массовый процесс – выселение нейробластов из эпендимного в мантийный слой. При формировании коры больших
полушарий головного мозга и мозжечка нейробласты
выселяются из эпендимного слоя в мантийный слой и
краевую вуаль. В мозжечне они формируют слой клеток Пуркинье. Важное значение в
направленной миграции клеток в пределах нервной трубки имеют специальные
поддерживающие клетки радиальной глии.
Направленный рост аксонов заключается в том, что нейрон «узнает» свою область
иннервации и находит своего клеточного
партнера и в развивающемся мозге многочисленные отростки нейронов
устанавливают связи с большой точностью.
Отросток нейрона (аксон) сразу и без ошибок находит свои мишени. Направленный
рост аксонов осуществляет конус роста. Конус роста - специализированная структура терминали растущего аксона имеет на конце булавовидное
утолщение (ламеллоподия), от которого отходят тонкие
пальцевидные отростки - филлоподии. Они растут в
различных направлениях и исследуют потенциальное пространство роста аксона.
Согласно представлению С.Рамона-и-Кахаля о хемотропизме, рост аксонов происходит по
градиенту концентрации специфических химических факторов, вырабатываемых в
мишенях.
Наиболее разработано представление о меченых
путях, которые образованы молекулами – метками, закономерно
распределенными в потециальном пространстве роста
аксона. По мере роста аксон последовательно считывает одну за другой метки,
расположенные в межклеточном пространстве или на поверхности клеток, и растет в
нужном направлении. Вслед за ним мигрируют отростки других аксонов, совокупность
которых формирует тракты в ЦНС и нервы на периферии
Ключевой момент
представления о заранее размеченных путях – узнавание – обеспечивают молекулы
адгезии, которые встроены в
плазмалемму ламеллоподии и филлоподий
и взаимодействуют с комплементарными молекулами (ламинин,
фибронектин, коллаген, тинасцин
и др.) во внеклеточном пространстве. Это обеспечивает фиксацию конуса роста на
поверхности мишени в нужном месте в нужное время.
Физиологическая гибель клеток (апоптоз) –
запрограммированная массовая смерть нейронов на определенных этапах онтогенеза.
Естественная гибель нейронов прослежена в ЦНС и периферической нервной системе.
Объем субпопуляции гибнущих нейронов оценивают в
пределах от 25% до 75%. Иногда в популяции погибают все нейроны. Выраженная
гибель нейронов в сформированной нервной ткани наблюдается при дегенеративных
заболеваниях нервной системы, а таких как болезнь Паркинсона и др.
Нейротрофические взаимодействия. Эта концепция предусматривает информационный обмен
между нейронами, отличающийся от химической передачи возбуждения в синапсах.
Нейротрофические взаимодействия реализуются через нейротрофические факторы. Эти
химические факторы вырабатываются в одних нейронах или ненервных клетках и
воздействуют на другие нейроны. Нейротрофические факторы обеспечивают выживание
и дифференцировку нейронов в нейроонтогенезе и при
регенерации. Наиболее изученным нейротрофическим фактором является фактор
роста нейронов.
Первые признаки дифференцировки нейробластов
выражаются в образовании фибриллей, с появления
которых можно говорить о формировании нейронов. Одновременно с дифференцировкой
фибриллей развиваются отростки.
Закладка аксона проявляется в виде булавовидного
выроста, который быстро растет. Дендриты закладываются позднее и, не вырастая
сильно в длину, начинают ветвиться вблизи перикариона.
С образованием отростков нейробласт превращается в
нейрон с одним аксоном и разным количеством дендритов, ориентированных в разных
направлениях.
Дальнейшее развитие нейробластов
ведет к удлинению отростков, которые образуют пучки нервных волокон, причем
рост аксонов идет быстрее, чем дендритов. Аксон в дальнейшем принимает участие
в образовании нервного волокна.
Вначале отростки лишены мякотных оболочек, клетки
нейроглии образуют их на более поздних стадиях эмбриогенеза. На 5-ом месяце
эмбрионального развития человека образуются синапсы, в клетках
увеличивается количество нейрофибрилл и начинают
возникать мякотные оболочки. С появлением этих структурных элементов
завершается дифференциация нейробластов в нейроны.
Интенсивный рост аксонов и дендритов продолжается и в
первые месяцы после рождения. В это время резко возрастает количество синапсов.
По пути следования отростков нервных клеток за пределы мозга смещаются и
некоторые нейроны, формирующие впоследствии периферические ганглии
симпатической и парасимпатической нервной системы.
У высших позвоночных и человека все нейробласты, появляющиеся в нервной закладке, превращаются
в нейроны. Поэтому в постнатальном онтогенезе не происходит образования новых
нейронов. Следовательно, погибшие нейроны не восстанавливаются. Из этого не
следует, что в нервной системе отсутствует
регенерация. Некоторые специализированные элементы (отростки нейрона)
обладают частичной способностью к восстановлению.
Долгое время считали, что популяция нейронов относится
к замкнутой статической. Это означает, что во
время постэмбрионального онтогенеза данная популяция не пополняется и клетки из
нее не переходят в следующую популяцию. Такие клетки сохраняются в течение
жизни особи и лишь единичные из них гибнут. Ткани,
относящиеся к статическим, очевидно, способны к своеобразной форме регенерации,
которая совершается не путем размножения клеток, а с помощью внутриклеточных
восстановительных процессов. Характерным свойством нейронов
является их способность к полиплоидизации.
Наряду с этим в последние годы показана возможность
возникновения новых нервных клеток в мозге взрослых млекопитающих. Впервые нейрогенез был открыт в 1962 году американским ученым Жозефом Олтманом, который в мозг крысы после разрушения
электротоком одной из его структур вводил радиоактивное вещество и через
несколько месяцев обнаружил в нем новые радиоактивные нейроны.
В середине 1980-х годов нейрогенез был обнаружен в мозге птиц. Американский
профессор из Рокфеллеровского университета Фернандо Ноттембум показал, что у взрослых самцов канареек процесс нейрогенеза происходит постоянно в вокальном центре, но
количество образующихся нейронов подвержено сезонным колебаниям.
В конце 1980-х годов нейрогенез
также был обнаружен у взрослых амфибий в лаборатории ленинградского ученого А.Л.Поленова. Источником новых нейронов у птиц и амфибий
оказались нейрональные стволовые клетки стенки желудочка
мозга. В начале 1990-х годов
возобновляемые нейроны были обнаружены в головном мозге взрослых крыс,
мышей и человека. Они были найдены во всех отделах головного мозга, включая
кору больших полушарий. Новые нейроны во взрослом мозге могут образовываться не
только из нейрональных стволовых, но и из стволовых
клеток крови. Оказалось, что стволовые
клетки крови действительно проникают в
мозг, но они не превращаются в нейроны,
а сливаются с ними, образуя двуядерные клетки. Затем ядро нейрона
разрушается и его замещает ядро стволовой клетки крови. Согласно одной из
гипотез, стволовые клетки крови несут с собой новый генетический материал,
который, попадая в клетки мозжечка, продлевает им жизнь.
Что же касается нейроглии, то она во
взрослом организме содержит малодифференцированные клетки, способные к
размножению и развитию в течение всей жизни. При повреждении центральных
отделов нервной системы в очаге повреждения скапливается много астроцитов, которые закрывают рану. Погибшие перикарионы нейронов и их отростки распадаются на отдельные
фрагменты и фагоцитируются клетками микроглии и, очевидно, астроцитами,
в цитоплазме которых обнаруживаются лизосомы.
Элементы сравнительной гистологии и
эволюции
нервной системы
Нервная система прошла длительную эволюцию в процессе
филогенетического развития животных. С повышением их организации происходило
усложнение нервной системы и форм ответных реакций на воздействия внешней
среды.
Развитие и совершенствование нервной системы в
процессе эволюции было тесно связано с дифференцировкой двигательных тканей. У
наиболее низкоорганизованных беспозвоночных - губок – имеются сократимые
клетки, но отсутствует нервные. А.А. Заварзин
рассматривал сократимые клетки как прообразы нервной и мышечной систем вместе
взятых.
Первичные чувствительные клетки появляются в эктодерме
и энтодерме кишечнополостных. Просто диффундированные
и рассеянные по всему телу, они образуют диффузную нервную систему. Диффузная
нервная система (нервные сплетения) представляют собой сеть из униполярных, биполярных
и мультиполярных клеток, связанных между собой.
Усложнение нервной системы шло в
направлении концентрации нервных элементов в определенных частях тела и привело
к образованию нервных центров, которые у примитивных животных расположены в нервных
стволах. У низших червей турбелярий нервная
система состоит из головного нервного узла и нервных тяжей, идущих вдоль тела.
Она имеет эктодермальное происхождение. В ходе
дальнейшей эволюции происходит скопление нервных клеток в нервных узлах (ганглиях),
которые образуют цепочечную (ганглиозную) нервную систему. Она характерна большинству беспозвоночных
животных. У аннелид ЦНС состоит из окологлоточного нервного кольца и
брюшной нервной цепочки, состоящей
из метамерных нервных узлов и парных комиссур. Окологлоточное нервное кольцо
организовано из надглоточных и подглоточных ганглиев,
связанных между собой комиссурами. От брюшной нервной цепочки отходят нервы к
различным органам.
Таким образом,
централизация нервной системы у высших представителей аннелид выражена весьма
значительно. Такие прогрессивные изменения в нервной системе происходили вместе
с усложнениями поведенческих реакций, требующих согласованной работы мышц и
нервной системы. У высших беспозвоночных происходит редукция нервных сплетений.
У моллюсков наряду с центральным нервным аппаратом,
состоящим из ганглиев, имеются и нервные сплетения напоминающие таковые у кишечнополостных. Особенно сильно процесс цефализации наблюдается у головоногих моллюсков. У высших
моллюсков централизация нервной системы хорошо выражена, хотя диффузные
сплетения все еще сохраняются.
Для членистоногих характерна полная редукция
подкожного нервного сплетения. У многих ракообразных, насекомых и у всех
хелицеровых происходит полное слияние ганглиев в одну непарную брюшную
нервную цепочку.
У иглокожих нервная система
построена по лучевой симметрии. Имеется нервное кольцо вокруг пищевода и
нервные стволы, расходящиеся по лучам от кольца. От стволов отходят нервы к амбулакральным ножкам.
У полухордовых
(низших вторичноротых) и бесчерепных (ланцетник) нервная система находится на низком уровне
организации. У них имеются нервные сплетения не только под эктодермой, но и
вокруг кишечника. Обособленного центрального отдела нервной системы нет. Центральный отдел нервной системы ланцетника
представлен спинномозговой трубкой, которая тянется вдоль тела. Диффузные
нервные сплетения у ланцетника отсутствуют, однако чувствительные нервные
клетки еще не сконцентрировались в спинальных ганглиях, а расположены вдоль
чувствительных нервов вдалеке от иннервируемого органа.
У позвоночных животных центральная и периферическая
нервная система достигает наивысшей степени развития. Вегетативная нервная
система бесчерепных и круглоротых еще содержит
ганглии. Вегетативные ганглии впервые появляются у акуловых,
у них же намечается головной (парасимпатический) брюшной (симпатический) отделы
вегетативной нервной системы.
У амфибий, рептилий, птиц и млекопитающих
вегетативная нервная система приобретает все большую сложность организации и
специфические структурные особенности.