Лекция 6.
Костные ткани.
Строение,
химический состав и значение костных тканей
Опорная функция у беспозвоночных
животных выполняется эпителиальными тканями. У них отсутствует скелет, типичный
для хордовых животных. У хордовых животных опорная функция переходит от
пограничных тканей (эпителия) к соединительным в
дополнение к их основной первичной функции обмену веществ. На основе этой
функции в филогенезе возникли скелетные ткани внутренней среды, из которых
наиболее типичны костные ткани.
Костные ткани имеются у всех
позвоночных животных, за исключением круглоротых и хрящевых рыб. Они выполняют
опорную функцию. Костные ткани формируют скелет организма человека и
позвоночных животных, определяют форму тела, защищают
и поддерживает жизненно важные органы, расположенные в черепе, грудной и
тазовой полостях, позвоночнике. Они играют важную роль в минеральном обмене
организма, выполняя функцию депо кальция. До 99% всего кальция, имеющегося в
организме, содержится в костных тканях. В костных тканях находится до 90% всей
содержащейся в организме человека лимонной кислоты.
Красный костный мозг является
кроветворным органом и выполняет защитную функцию, т.к. в нем возникают
макрофаги. В полостях трубчатых костей, в желтом костном мозге накапливается
большое количество жира, принимающий участие в жировом
обмене.
Костные ткани относятся к разновидностям
тканей внутренней среды с ярко выраженной механической функцией. Они – из самых
твердых соединений. По твердости они уступают лишь эмали зубов. Костные ткани
состоят их клеток и минерализованного
матрикса. Костный матрикс составляет 50% сухой массы кости и
содержит неорганические (50%0 и органические (25%) вещества и 25% воды..
Неорганическая часть больше всего содержит кальций (35%) и фосфор (50%),
образующие комплексную соль в виде кристаллов гидроксоапатита.
Кристаллы гидроксоапатита соединяются с молекулами
коллагена через остеонектин. В состав неорганической
части кости также входят бикарбонаты, цитраты, фториды, соли Mg+2, K+, Na+. С возрастом количество неорганических солей в костях
увеличивается, поэтому кости старых людей становятся хрупкими и легко
подвергаются переломам.
Органическая
часть кости составляют коллаген
типа І (90-95%) и типа V, а также неколлагеновые белки (остеонектин,
остеокальцин, протеогликаны,
сиалопротеиды, морфогенетические белки, протеолипиды, фосфопротеиды, а
также гликозаминогликаны, (хондроитинсульфат,
кератинсульфат). Органические вещества костного матрикса синтезируют остеобласты. Остеонектин участвует в процессе кальцификации,
служит маркером для оценки активности костной ткани. Морфогенетические белки
костной ткани – регуляторные, они индуцируют энхондральный
остеогенез.
В целом органические
вещества придают упругость костям, минеральные соли – твердость. Сочетание
твердости и упругости обусловливает большую прочность костей.
К клеткам костных тканей
относятся остеобласты, остеоциты и остеокласты. В кости присутствуют две линии
клеток - созидающая и разрушающая, что отражает постоянство происходящий
процесс перестройки костной ткани. К созидающей линии клеток относятся
остеобласты и остеоциты. Разрушающая линия клеток – остеоклас
Остеобласты – клетки, образующие костную ткань. Они неделящиеся отростчатые клетки. Активно синтезируют и секретируют
вещества костного матрикса. Они встречаются в участках, где происходит
перестройка костной ткани.
Остеобласты – клетки, образующие костные ткани. Они неделящиеся отростчатые клетки. Активно синтезируют и секретируют
вещества костного матрикса. Они встречаются в участках, где происходит
перестройка костной ткани. В эмбриональной кости остеобласты покрывают всю
поверхность оссеидной ткани. Они имеют цилиндрическую
или угловатую форму, короткие отростки, которыми соприкасаются друг с другом.
Клетка имеет хорошо развитую гранулярную ЭПС, комплекс
Гольджи, митохондрии, множество секреторных гранул.
Характерным свойством остеобластов является высокая активность щелочной
фосфатазы, играющей важную роль в минеральном обмене кости.
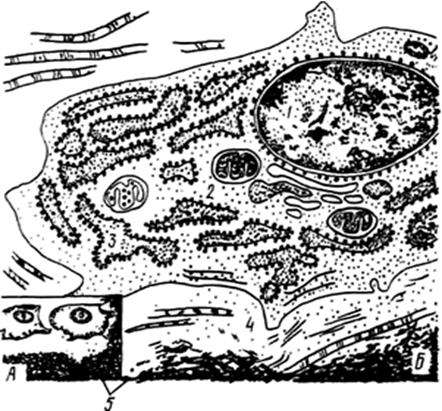
Рис.25. Схема строения остеоблста:
А – на светооптическом; Б – на субмикроскопическом
уровне; 1 – ядро; 2 – цитоплазма; 3 –гранулярная
эндоплазматическая сеть; 4 – остеоид; 5 – минерализованное вещество костной ткани.
Остеоциты – основные клетки костных тканей, образующиеся из
остеобластов. Остеоциты имеют звездчатую форму и отростки, соприкасающиеся с
отростками соседних клеток. Ядро округлое или овальное.
 Рис.26.
Костные клетки (остеоцит):
Рис.26.
Костные клетки (остеоцит):
1 – ядро; 2 – цитоплазма; 3 – отростки.
Гранулярная ЭПС и цистерны комплекса Гольджи
развиты слабее, чем в остеобластах. Остеоциты
поддерживают структурную целостность минерализованного матрикса, участвуют в регуляции
обмена Са2+ в организме. Эта функция остеоцитов находится
под контролем со стороны Са2+ плазмы
крови и различных гормонов. Остеоциты могут секретировать вещества для
образования матрикса новой кости, но эта способность менее выражены, чем у
остеобластов. Остеоциты – зрелые неделящиеся клетки. Они находятся в костных
полостях, которые называются лакунами.
Совокупность сообщающихся между собой канальцев и лакун составляет лакунарно-канальцевую систему, заполненную тканевой
жидкостью, через которую осуществляется обмен веществ между остеоцитами и
кровью. 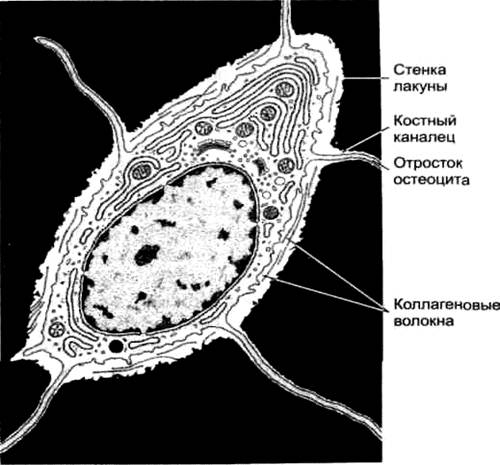
Рис.27. Остеоцит. Тонкие длинные отростки
проходят в костных канальцах. Междустенкой лакуны и
поверхностью остеоцита в составе минерализованного матрикса (остеоида) расположены
коллагеновые волокна.
В канальцах постоянно циркулирует жидкость, благодаря
чему поддерживается диффузия метаболитов и обмен между лакунами и кровеносными
сосудами надкостницы. По химическому составу лакунарно – канальцевая
жидкость отличается от плазмы крови.
Разделяющий плазму и лакунарно-канальцевую
жидкость барьер называется костной мембраной. Барьер формируют
остеобласты и остеоциты. Концентрация Са+2 и РО-34
в лакунарно-канальцевой жидкости превышает
критический уровень, что способствует спонтанному осаждению солей кальция.
 Рис.28.
Схема строения остеокласта:
Рис.28.
Схема строения остеокласта:
А – на светооптическом уровне; Б – на субмикроскопическом уровне; 1 – ядра; 2 –
гофрированный край остеокласта; 3 –
светлая зона; 4 – лизосомы; 5 – зона резорбции межклеточного
вещества; 6 – минерализованное
межклеточное вещество.
Остеокласты – «костедробители», клетки,
разрушающие кость. Они участвуют в обновлении костной ткани. Остеокласты –
крупные неклеточные структуры – симпласты, достигающие размеров до 100мкм и содержащие
много ядер (до 20 и более). Они имеют ацидофильную цитоплазму и содержат гидролитические
ферменты. Остеокласты образуются путем слияния моноцитов. Остеокласты относятся
к системе мононуклеарных фагоцитов. Для дифференцировки остеокластов необходим колониестимулирующий
фактор макрофагов (M-CSF) и остеопротегерин лиганд (OPGL),
продуцируемые ретикулярными клетками костного мозга и остеобластами.
Остеокласт прикрепляется к резорбируемой
поверхности кости за счет формирования замыкающего кольца из подосом – временных выростов цитоплазмы светлой зоны,
соединяющих F-актин, винкулин,
талин, α-актинин.
Кольцо подосом исчезает после завершения резорбции и
ухода остеокласта. Через мембрану выростов из остеокласта выделяется большое
количество Н+ и Cl-, что
создает и поддерживает в замкнутом пространстве лакуны кислую среду,
оптимальную для растворения солей кальция костного матрикса.
Образование Н+ в цитоплазме остеокласта
катализирует карбоангидраза II:
карбоангидразаII
СО2 + Н2О
↔↔Н+ + НСО3-
Н+
при помощи протонной Н+,К+ -АТФ
- азы активно выкачивается из клетки, что приводит к закислению
замкнутого пространства лакуны.
Везикулярная зона цитоплазмы остеокласта содержит
лизосомы, ферменты которых (кислые гидролазы, коллагеназы,
катепсин К) разрушают органическую часть костного
матрикса.
Классификация костных тканей
По структуре костного матрикса и в зависимости от
расположения коллагеновых волокон костные ткани подразделяются на 2 вида: ретикулофиброзную (грубоволокнистую, первичную, незрелую) и
пластинчатую (вторичную, зрелую).
Грубоволокнистая (первичная, или незрелая) костная ткань образован
толстыми пучками коллагеновых волокон, расположенных в виде сети. В основном веществе находятся костные лакуны
с длинными канальцами. В лакунах находятся остеоциты с отростками. Характерно большое количество протеогликаов и гликопротеидов и низкое содержание
минеральных солей.
Из грубоволокнистой костной ткани построен
скелет низших позвоночных (рыб, амфибий) и скелет зародышей высших позвоночных.
У взрослых особей она встречается лишь в местах прикрепления сухожилий костям,
вблизи черепных швов, в зубных альвеолах, костном лабиринте внутреннего уха.
Постнатально незрелая кость часто образуется при заживлении переломов и в
быстро растущих костных опухолях, возникающих из остеогенных клеток.
У высших позвоночных скелет образован пластинчатой
(зрелой, или вторичной) костной тканью. Она образована
костными пластинками. Костные пластинки образованы
упорядоченно расположены минерализованными коллагеновыми волокнами.
Костная
пластинка – структурная единица пластинчатой костной ткани. Длина и
толщина костной пластинки варьируют от нескольких десятков до сотен мкм. Между соседними пластинками в лакунах расположены
остеоциты, а в толще пластинки в костных канальцах проходят их отростки.
Коллагеновые волокна в пределах пластинки расположены параллельно и лежат под
углом к волокнам соседней пластинки, что обеспечивает значительную прочность
пластинчатой кости. Пластинчатая костная ткань больше минерализована, чем
грубоволокнистая
Пластинчатая костная ткань формирует
губчатое и компактное вещество кости. Соответственно этому различают
пластинчатую губчатую и пластинчатую компактную костную ткань.
Пластинчатая губчатая костная ткань образована переплетающимися костными трабекулами,
полости между которыми заполнены костным мозгом. Трабекула состоит из костных
пластинок и снаружи окружена одним слоем остеобластов. Трабекулы расположены
соответственно направлению сил сжатия и растяжения. В губчатом веществе
отсутствуют остеоны. Губчатое вещество заполняет эпифизы трубчатых костей и
образует внутреннее содержимое коротких и плоских костей скелета.
Распределение губчатого компактного
вещества в различных костях определяется функциональным значением данной кости,
условиями давления и натяжения. В зависимости от характера и степени нагрузки
структура губчатого вещества изменяется в пользу компактного вещества при
увеличении нагрузки (например, ногтевые фаланги у балерин).
Пластинчатая компактная костная ткань состоит из костных пластинок, которые тесно прилегают друг к другу. Из
нее состоят диафизы трубчатых костей и слоем различной
толщины покрывает все остальные короткие и плоские кости скелета.
 Структурной-функциональной единицей является гаверсова
система(остеон) – система костных пластинок, в
которой расположен центральный, или гаверсов, канал. В гаверсовом канале проходят
кровеносные сосуды и нервы и он заполнен рыхлой
соединительной тканью. Вокруг них костные пластинки расположены правильными
кругами концентрически (по 4-20), образуя как бы цилиндры, вставленные один в
другой. Между ними лежат костные лакуны с остеоцитами.
Структурной-функциональной единицей является гаверсова
система(остеон) – система костных пластинок, в
которой расположен центральный, или гаверсов, канал. В гаверсовом канале проходят
кровеносные сосуды и нервы и он заполнен рыхлой
соединительной тканью. Вокруг них костные пластинки расположены правильными
кругами концентрически (по 4-20), образуя как бы цилиндры, вставленные один в
другой. Между ними лежат костные лакуны с остеоцитами.
Рис.29 .
Пластинчатая кость:
А – плотное (компактное) вещество кости; 1 –
надкостница; 2 – наружные общие пластинки; 3 – остеоны; а –
канал остеона; 4 – система вставочных пластинок; 5 – внутренние
общие пластинки; Б – губчатое вещество кости; 6 – желтый костный
мозг.
Костные пластинки имеют толщину 4,5-12мкм и состоят из
оссеомукоида и параллельно расположенных коллагеновых
волокон, импрегнированных кристаллами гидроксоапатита.
Вся система пластинок с гаверсовым каналом в середине называется гаверсовой
системой, или остеоном.
Диаметр остеона (не более
В ходе образования остеона находящиеся в
непосредственной близости от сосуда гаверсова канала остеогенные клетки
дифференцируются в остеобласты. Снаружи располагается сформированный
остеобластами слой остеоида. В дальнейшем остеоид минерализуется и остеобласты, окруженные
минерализованным костным матриксом, дифференцируются в остеоциты. Следующий
концентрический слой возникает подобным же образом изнутри. По наружной
поверхности остеоида на границе с минерализованным
костным матриксом проходит фронт обызвествления, где начинается процесс
отложения минеральных солей.
Трубчатая кость состоит из большого количества
остеонов, которые расположены вдоль ее длинной оси. В плоских костях остеоны
идут параллельно их поверхности, в телах позвонков – перпендикулярно оси.
Благодаря такому строению пластинчатая кость достигает значительной
механической прочности.
Остеоны расположены на некотором
расстоянии друг от друга. Между остеонами находятся вставочные костные пластинки.
С наружной поверхности трубчатые кости покрыты системой наружных генеральных пластинок.
Внутренние поверхности костных полостей выстланы внутренними генеральными
пластинками. Со стороны внутренних генеральных пластинок кость
выстлана соединительнотканной оболочкой – эндостом.
Эндост – тонкая оболочка, покрывающая трабекулы в губчатом
веществе, а также выстилающая кость (со стороны костного мозга) и гаверсовы
каналы компактного вещества. Эндост присутствует на поверхности всех костных полостей.
Он состоит из слоя неактивных плоских остеогенных клеток. В период роста и
перестройки кости целостность эндоста часто
нарушается остеокластами.
Снаружи вся кость, за исключением
суставных поверхностей, покрыта надкостницей (периостом). Периост
состоит их двух слоев: наружного – волокнистого и внутреннего - остеогенного.
Наружный волокнистый слой надкостницы более
плотный, состоит из толстых пучков коллагеновых волокон, которые обусловливают
их прочность. Он образован плотной волокнистой соединительной тканью. В этом
слое проходят нервы и кровеносные сосуды. Отсюда они проникают в гаверсовы
каналы и питают кость. Пучки прободающих коллагеновых волокон (волокна Шарпея) и уходящие в матрикс кости из надкостницы
обеспечивают прочное прикрепление внутреннего слоя к поверхности кости.В таких местах надкостницу
трудно отделить от кости. Волокнистый слой надкостницы защищает остеогенные
клетки от механических повреждений.
Внутренний остеогенный слой содержит рыхлую
волокнистую ткань, кровеносные сосуды и остеогенные клетки, обеспечивающие
аппозиционный рост и регенерацию костей.
Костный мозг. Полости всех костей эмбриона заполняет красный костный мозг - орган
кроветворения. Он образован ретикулярной тканью, в петлях которой расположены
кровяные клетки, находящиеся на разных стадиях развития. После рождения ребенка
красный костный мозг остается лишь в эпифизах трубчатых костей, в телах
позвонков и в некоторых плоских костях. В остальных костях он замещается желтым
костным мозгом. Он состоит из жировых клеток, образующихся в результате
накопления жира в ретикулярных элементах, и в кроветворении не участвует.
Развитие костей (остеогенез)
Различают внутримембранный
(прямой) и энхондральный (непрямой) остеогенез. При внутримембранном остеогенезе кость развивается непосредственно из
мезенхимы (кости черепной крышки и лица). При энхондральном остеогенезе кости развиваются на месте гиалинового
хряща (позвонки, кости конечностей и их поясов, основание черепа). В обоих
случаях сущность костеобразования одинакова и источником костной ткани служит
мезенхима. В обоих случаях вначале образуется грубоволокнистая, а позже
пластинчатая костная ткань.
Внутримембранный (прямой) остеогенез. Этим способом у млекопитающих и человека образуются
плоские кости – кости лицевого отдела черепа и покровных костей мозговой
коробки черепа. При образовании костной ткани из мезенхимы наблюдается
несколько стадий.
На I-ой стадии происходит формирование остеогенного
островка. Определенные участки мезенхимы, содержащие капилляры, уплотняются за
счет размножения клеток и происходит васкуляризация этих островков.
На II-ой стадии группы мезенхимных клеток формируют
первичные центры окостенения. Мезенхимные клетки дифференцируются в
остеобласты, которые начинают вырабатывать коллагеновые волокна типа I, образующие основоу остеоида. В аморфном веществе образуются остеомукоиды, которые склеивают волокна в прочную массу.
Таким образом клетки оказываются погруженными в
межклеточное вещество остеоида. В таком виде она
отличается от окружающей мезенхимы и называется скелетогенной закладкой. Костная закладка растет за счет дифференцировки
остеобластов.
III-ая стадия – оссификация (обызвествление) остеогенного вещества. Остеобласты выделяют фермент щелочную фосфатазу,
которая расщепляет глицерофосфаты крови. Образуется Н3РО4,
которая вступает в реакцию с СаCL2 с
образованием Са3(РО4)2.
Фосфат кальция вместе с СаСО3
обызвествляет основное вещество и волокна –
образуется основное костное вещество. Остеонектин связывает соли кальция и фосфора с
коллагеном. Мезенхима вокруг формирующейся кости образует надкостницу.
Таким
путем остеоид минерализуется
и дифференцирующиеся остеоциты оказываются замурованными в лакунах
минерализованного костного матрикса. На этой стадии образуется грубоволокнистая
(первичная) костная ткань. Сформировавшаяся незрелая костная ткань существует в
форме трабекулы. Отдельные трабекулы, образовавшиеся в различных участках,
растут и объединяются друг с другом. Анастомозирующаяся
сеть костных трабекул формирует губчатое вещество. Поверхность трабекул
покрывает слой остеобластов и остеогенных клеток. За счет этого слоя происходит
аппозиционный рост с образованием костных пластинок. Оно связано с постепенным
разрушением первичной кости остеокластами и ее замещением зрелой пластинчатой
костной тканью. Это IV-ая
стадия остеогенеза. Эта перестройка происходит под
влиянием деятельности двух видов клеток: разрушающих кость – остеокластов и
образующих ее – остеобластов.
Энхондральный (непрямой) остеогенез происходит на месте гиалинового хряща и имеет место как в эмбриональном, так и в постэмбриональном периоде
развития.
 Рис. 30. Развитие пластинчатой костной ткани.
Рис. 30. Развитие пластинчатой костной ткани.
1 – распадающаяся хрящевая
клетка, 2 – перихондральная
кость, 3 – остеоцит; 4 – надкостница, 5 – обызвествляющийся хрящ, 6 – энхондральная кость, 7 – остеобласт, 8 – сосуд; 9 – остеокласт
По этому
типу развиваются трубчатые кости скелета. Образование кости на месте хряща
совершается сложнее, чем непосредственно из мезенхимы. Вначале у эмбриона
возникает хрящевая модель будущей кости, которая состоит из гиалинового хряща.
Хрящевая модель служит основой для развития кости. В развивающейся костной закладке параллельно идут два
процесса окостенения: перихондральный – на
поверхности хрящевой модели и энхондральный внутри нее, на месте разрушающего
хряща. При этом хондроциты экспрессируют
редкий тип (Х-й) коллагена. Морфогенетические белки кости индуцируют энхондральный остеогенез.
Выделяют
несколько стадий энхондрального остеогенеза:
1) образование хрящевой модели кости;
2) формирование перихондральной
костной манжетки;
3) развитие энхондральной кости
в диафизе;
4) развитие энходральной кости
в эпифизе;
5) окостенение диафиза и эпифиза и формирование эпифизарной пластинки роста в хряще;
6) замещение эпифизарной
пластинки роста костью.
Первоначально
из скелетогенной мезенхимы образуется эмбриональный хрящ, который является гиалиновым.
Она имеет надхрящницу. У млекопитающих окостенение начинается с центральной
части диафиза хряща. В надкостницу
проникает кровеносные сосуды и скелетогенные клетки надхрящницы, которые
начинают дифференцироваться в остеогенные (остеобласты). Надхрящница
превращается в надкостницу. Остеобласты начинают синтезировать межклеточное
вещество костной ткани и поверх хряща создают костную ткань в виде манжетки (перихондральное окостенение).
В
диафизе первичная костная ткань замещается компактным веществом. Образованная ранее костная манжетка
утолщается и растет по направлению от диафиза к эпифизам. Вслед за диафизом
центры окостенения появляются и в эпифизах.
В эпифизах оссификация протекает аналогично формированию диафизарного центра окостенения, но на месте первичной костной ткани образуется
губчатое вещество. Оссификация начинается с врастания
в эпифиз кровеносных сосудов. Когда новообразованная костная ткань заполнит
весь эпифиз, хрящевая ткань остается в виде узких полосок только на поверхности
эпифиза (суставной хрящ), а также между эпифизом и диафизом в виде эпифизарной хрящевой пластинки. Эпифизарная хрящевая пластинка не окостеневает у человека
до 23-25 лет (у женщин раньше) и за счет ее кости растут в длину. В дальнейшем процесс
образования костной ткани в зоне эпифизарной линии
тормозится, хрящ замещается костной тканью и рост
кости в длину прекращается. В толщину кость растет до преклонного возраста
организма, поскольку рост ее идет за счет остеонов и надкостницы. Рост
трубчатых костей в ширину происходит благодаря образованию новых слоев костной
ткани (образующихся путем аппозиционного роста) остеогенными клетками
надкостницы. По мере завершения роста кости под надкостницей формируются
наружные генеральные пластинки (также по аппозиционному механизму). Все эти
процессы имеют существенное значение для формирования костей
и нарушение даже одного из них приводит к атипичному остеогенезу
и ненормальному росту костей.
Влияние условий внутренней и
внешней среды
на развитие
костей
Кость
как сложный орган подвергается постоянному развитию и обновлению. Интенсивность
этих процессов зависит от внутренних и внешних факторов. К внутренним факторам
следует отнести гормональные влияния половых, околощитовидных и щитовидной
желез, передней доли гипофиза, надпочечников, воздействие ЦНС. К внешним
факторам относятся факторы питания, особенно содержание соединений кальция и
фосфора, витаминов А, С, Д, а также солнечное
облучение (УФ–спектр).
Для нормальной
минерализации остеоида особенно необходим
1α, 25-дигидроксихолекальциферол (активная форма витамина Д3 – кальцитриол). Способствуя всасыванию кальция и фосфора в
кишечнике, кальцитриол
обеспечивает их необходимую концентрацию для запуска кристаллизационных
процессов в костном матриксе. Прямо воздействуя на
остеобласты,1α,25-дигидроксихолекальциферол повышает активность щелочной
фосфатазы в этих клетках, способствуя минерализации костного матрикса.
Обмен кальция и
фосфатов в организме регулируют гормон паращитовидной железы и кальцитонин.
Гликокортикоиды ухудшают всасывание Са+2 в кишечнике. Транспорт кальция в
ЖКТ в значительной мере определяет кальцитриол.
Паратиреоидный гормон повышает содержание кальция в крови за счет увеличения
его высвобождения из костей и снижения его экскреции почками.
Кальцитонин – пептидный гормон, синтезируемый С-клетками
щитовидной железы, снижает уровень кальция в крови за счет ингибирования
резорбции костей остеокластами. Формирование кости и
ее минерализация стимулируют соматотропин и половые
гормоны. При недостатке эстрогенов развивается остеопороз.
Витамины А и С имеют важное
значение для роста костей. Витамин С необходим для
образования коллагеновых волокон. При дефиците этого витамина замедляется рост
костей и заживление переломов. Витамин А поддерживает
образование и рост кости. Недостаток витамина А
тормозит остеогенез и рост костей. Кальцитриол, необходимый для всасывания Са2+ в тонкой кишке, поддерживает
процесс минерализации костей. Дефицит витамина Д
приводит к нарушению минерализации
костей, что и наблюдается при рахите у детей и остеомаляции у взрослых.
Синтез макромолекул костного матрикса
стимулируют соматомедины.
Регенерация
костей
В течение всей жизни
организма кость все время перестраивается, происходит ее обновление. В костных
тканях постоянно протекают процессы резорбции старой кости и формирования
новой. Кость – динамичная структура с постоянно изменяющейся формой и
внутренней организацией. Происходит постоянная перестройка костной ткани, адаптирующая
кость к механическим нагрузкам.
Регенерация костей, как
и органов, после их переломов связана с деятельностью надкостницы. Кости, как
правило, хорошо регенерируют. Однако некоторые кости (кости свода черепа) плохо
регенерируют. Для успешной регенерации костей черепа оказывает благоприятное
влияние внесение в область повреждения костных опилок.
Известно,
что переломленные кости человека срастаются. Если между отломленными концами
нет соединительнотканных прослоек и расстояния между ними незначительные
(несколько мм), то кость может полностью восстановить свою целостность. При
этом регенерация кости происходит по механизму остеогенеза.
Если умело свести их части, то восстановление происходит без дефекта. В области
перелома повреждены ткани, нарушено кровоснабжение и
остеоциты в прилегающих участках остеонов гибнут. Отмирающая кость подвергается
активной резорбции. Между концами обломков формируется новая ткань – костная
мозоль. Костная мозоль возникает в результате интенсивного размножения
остеогенных клеток надкостницы. Часть этих клеток дифференцируется в
остеобласты, образующие новые костные трабекулы, прочно прикрепляющиеся к
матриксу обломка. Сначала формируется остеоид,
который затем минерализуется и формируется
грубоволокнистая костная ткань. Затем она замещается пластинчатой костной
тканью.
В случае, если скорость размножения остеогенных клеток в наружной
части костной мозоли превышает темпы роста кровеносных сосудов и
микроциркуляция в зоне перелома восстанавливается медленно, то дифференцировка
матричных клеток происходит в направлении образования хондробластов,
которые образуют хондрогенные островки и надхрящницу.
На месте перелома образуется хрящевая мазоль, состоящая из гиалинового хряща. В дальнейшем по
мере изменения микроциркуляции и формирования остеобластов хрящ замещается
грубоволокнистой костной тканью, на месте которой формируется губчатое
вещество. После этого костная мозоль перестраивается:
губчатое вещество между отломками преобразуется в компактное и
восстанавливается первоначальная конфигурация кости.
При простом закрытом
переломе, правильном совмещении отломков, при обездвиживании конечности
наступает сращение, т.е. восстанавливается структура кости в средней части
перелома, а по поверхности кости, на месте мозоли, остается след в виде
уплотнения, состоящего из неправильно отложенных наружных окружающих пластинок.
Кости конечности при
закрытом переломе требуют активного вмешательства хирурга для совмещения
отломков (консолидация) и их фиксирования иммобилизацией (лишение подвижности)
самой конечности. Это достигается наложением гипсовой повязки по Пирогову или
другими способами (по Илизарову).
Исследования
доказали на возможность активного вмешательства в процессы регенерации кости.
Существенную роль при этом играет пьезоэлектрический эффект. Многие
кристаллы обладают пьезоэлектрическими свойствами. При механической нагрузке
в них возникает электрический ток. Кость обладает выраженными свойствами
кристалла за счет гидроксоапатита и коллагена, находящегося в костном матриксе. Источник
электричества – точки соприкосновения кристаллов гидроксоапатита
и коллагена. В местах стыка этих двух веществ при
деформации (изгибе) кости появляется электрический потенциал. На перестройку,
регенерацию и трофику костной ткани оказывают воздействие биопотенциалы.
Остеобласты и остеоциты чувствительны к пьезоэлектрическим токам, возникающим
при деформации кости. Вогнутая поверхность кости несет отрицательный, а
выпуклая – положительный заряды. При отрицательном заряде в костной пластинке
наблюдается стимуляция остеобластов и аппозиционный рост, а при положительном
заряде - активация остеокластов и, как следствие, резорбция костной ткани.
Помимо частичной
наблюдается и полная регенерация костной ткани. Например, у хвостатых амфибий
при восстановлении отрезанных конечностей развиваются типичные для них кости. Полная регенерация конечностей
описана также для птиц и млекопитающих. Опыты показали восстановление трубчатых
костей петуха и некоторых млекопитающих. Во всех этих случаях необходимым
условием регенерации является сохранение надкостницы – только за счет ее
элементов развивается новая кость.