Лекция 3.
Кроветворение
Кроветворение (гемоцитопоэз,
гемопоэз) - это образование клеток крови,
происходящее в кроветворных органах. Различают эмбриональный и
постэмбриональный гемопоэз.
Эмбриональный
гемопоэз. В
эмбриональном
периоде развития кровь формируется как ткань. Эмбриональное кроветворение
характеризуется тем, что вместе с развитием крови происходит формирование и
сосудистой системы. В пренатальный период развития
человека кроветворение начинается с 3-й недели развития и может осуществляться
в различных органах развивающегося организма: в стенке желточного мешка,
печени, селезенке, тимусе и костном мозге. Соответственно этому последовательно
различают мегабластическую, гепатоспленотимическую
стадии и костномозговое кроветворение.
Мегабластическая стадия. Исходными клетками для возникновения всех клеток крови являются клетки
мезенхимы. Во внезародышевой мезодерме желточного мешка в течение 3-й недели
происходит формирование скоплений мезенхимных клеток – это кровяные островки.
Клетки, расположенные по периферии островка, дифференцируются в эндотелиальные
клетки первичных кровеносных сосудов. Эндотелиальные клетки пролиферируют и
образуют островки, в результате чего отдельные островки объединяются и
формируются первые мелкие сосуды. В центральной части островка образуются
первые клетки крови - первичные эритробласты,
содержащие ядро и эмбриональный гемоглобин. Лейкоцитов и тромбоцитов на этой
стадии еще нет. Кроветворение в желточном мешке завершается на 12-й неделе
развития зародыша человека.
Гепатоспленотимическая стадия начинается
на 2-м месяце эмбрионального развития, когда стволовые кроветворные клетки
заселяют печень, селезенку и тимус. В печени кроветворение начинается на
5-6-й неделе развития эмбриона. Здесь образуются эритроциты, гранулоциты и
тромбоциты. К концу 5-го месяца интенсивность гемопоэза
в печени снижается, но слабый процесс кроветворения продолжает идти еще
несколько недель.
Гемопоэз в селезенке наиболее выражен с 4-го по 8-й месяцы внутриутробного развития. В
селезенке образуются эритроциты, небольшое количество гранулоцитов и
тромбоцитов. Непосредственно перед рождением важнейшей функций селезенки
становится образование лимфоцитов.
В вилочковой железе первые лимфоциты
появляются на 7-8-й неделе развития зародыша. В течение 5-го месяца развития гемопоэз начинается в костном мозге, а к 7-му месяцу
костный мозг становится главным органом гемопоэза.
После рождения и до полового созревания
количество очагов кроветворения в костном мозге уменьшается. У взрослого
человека кроветворение ограничивается костным мозгом и лимфоидной тканью. Когда
костный мозг не в состоянии удовлетворить повышенный и длительный запрос на
образование клеток крови, гемопоэтическая активность печени, селезенки и
лимфатических узлов может восстановиться.
При эмбриональном гемопоэзе из клеток мезенхимы формируются первичные
кровяные клетки (гемоцитобласты), которые являются
малодифференцированными и могут превращаться в любые клетки крови. В связи с
этим их считают исходными, или стволовыми кроветворными клетками. В процессе развития стволовые кроветворные клетки мигрируют из желточного
мешка по кровеносному руслу сначала в печень, а затем костный мозг.
В ходе эмбрионального кроветворения
развитие форменных элементов крови начинается с образования эритроцитов,
которое осуществляется в сосудах стенки желточного мешка. Развитие лейкоцитов в
эмбриональном периоде начинается с образования гранулоцитов в печени, а затем в
красном костном мозге.
Постэмбриональное
кроветворение
Кроветворение во взрослом организме
млекопитающих и человека является процессом физиологической регенерации крови.
Установлено, что все форменные элементы крови происходят из единой стволовой
кроветворной клетки, популяция которой существует в течение всей жизни организма.
Стволовые кроветворные клетки млекопитающих и человека образуются в красном
костном мозге.
Красный
костный мозг содержит в большом количестве созревающие эритроциты, что придает
костномозговым очагам гемопоэза красный цвет. Строма
состоит из ретикулярных клеток с длинными отростками, ретикулиновых волокон, синусоидных капилляров и адипоцитов,
составляющих почти половину объема костного мозга. В полостях между
ретикулиновыми волокнами и отростками ретикулярных клеток расположены полости,
заполненные островками гемопоэтических клеток.
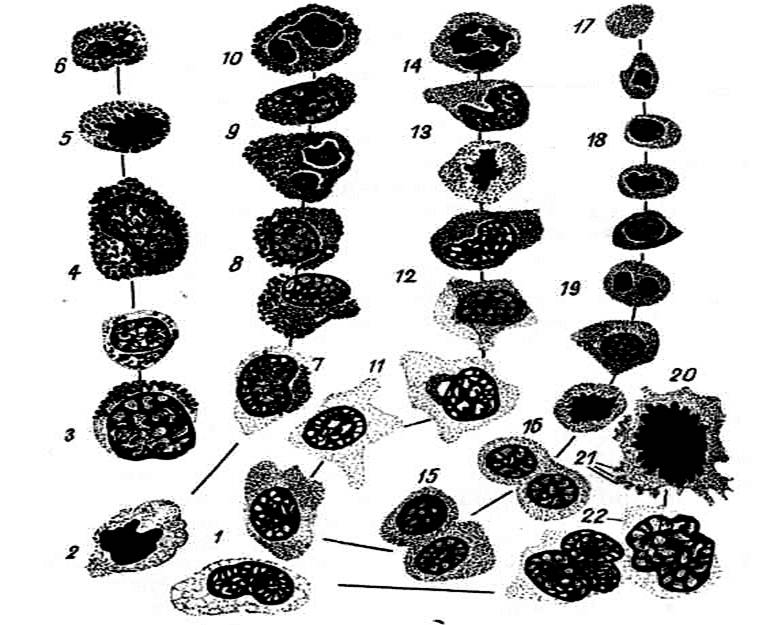
Рис.16. Различные стадии развития форменных элементов крови в красном
костном мозге:
1– гемоцитобласт
(стволовая клетка); 2 – митоз стволовой клетки; 3 –
базофильный промиелоцит;
4 – базофильный миелоцит; 5 –
базофильный метамиелоцит;
6 – базофильный гранулоцит; 7 – ацидофильный промиелоцит;
8 – ацидофильный миелоцит; 9
– ацидофильный метамиелоцит; 10 – эозинофильный гранулоцит; 11 – нейтрофильный промиелоцит; 12 – нейтрофильный
миелоцит; 13 – нейтрофильный метамиелоцит; 14 – нейтрофильный
гранулоцит; 15 – проэритробласт;
16 – базофильный эритробласт;
17 – эритроцит; 18 –
нормобласт; 19 – полихроматофильный
эритробласт; 20
– митоз мегакариоцита; 21 – тромбоциты; 22 –мегакариоцит
В костном мозге содержится большое количество макрофагов,
расположенных рядом с синусоидами. Помимо кроветворения в костном мозге, как и
в селезенке и печени, происходит удаление из кровотока старых и дефектных
клеток крови. Костный мозг играет центральную роль в иммунной защите, т.к. в
нем образуются В-лимфоциты, также присутствует большое количество
плазматических клеток, синтезирующих антитела. Образование Т-
и В-лимфоцитов может происходить и в лимфоидной ткани. Лимфоидная ткань входит
в состав органов иммунной системы - тимуса, селезенки, лимфатических узлов,
миндалин, червеобразного отростка.
У взрослого организма большая часть костного мозга
становится неактивной; в нем преобладают жировые клетки. Желтый костный мозг
может восстановить свою активность, если необходимо усилить гемопоэз
(например, при хронической гипоксии или выраженных кровотечениях).
В кроветворных органах клетки крови интенсивно
размножаются и находятся на различных стадиях зрелости. Созревшие форменные
элементы крови поступают в кровяное или лимфатическое русло и включаются в
циркуляцию. Развитие клеток крови идет таким образом, что около 60% стволовых
кроветворных клеток дифференцируется в эритроциты, около 30% - в лейкоциты и
около 5% - в мегакариоциты. Установлено, что Т-лимфоциты, воздействуя на
кроветворные органы, определяют направление клеточной дифференцировки при гемопоэзе. Таким образом, взаимодействие Т-лимфоцитов с
кроветворными стволовыми клетками является одним из механизмов регуляции
кроветворения.
Эритроцитопоэз. Начало эритроидного ряда
единица эритропоэза – эритропоэтинчувствительная клетка,
берущая свое начало от стволовой кроветворной клетки. На дальнейших стадиях эритропоэза дифференцируются проэритробласты,
эритробласты, ретикулоциты
и эритроциты. Длительность эритропоэза контролирует эритропоэтин. Основной стимул для выработки эритропэтина - гипоксия в тканях, зависящая от количества
циркулирующих эритроцитов Эритропоэтин усиливает
пролиферацию эритропоэтинчувствительных клеток в
костном мозге, что приводит к увеличению количества образующихся эритроцитов и,
как следствие, росту О2 в тканях. усилению
Униполярная колониеобразующая единица эритропоэза (эритропоэтинчувствительная
клетка) дает начало проэритробласту. Дальнейшая
дифференцировка приводит к уменьшению размеров клеток и количества органелл,
увеличению содержания гемоглобина и потере ядра. При этом из проэритобласта последовательно дифференцируются базофильный, полихроматофильный,
оксифильный эритробласт, ретикулоцит, эритроцит. От стадии эритропоэтинчувствительной
клетки до зрелых клеток проходит 6 последовательных делений.
Проэритробласты – крупные клетки, достигающие в диаметре 14-19 мкм,
лишенные гемоглобина. Ядро расположено эксцентрично, в цитоплазме много
рибосом. Проэритробласты многократно делятся митозом.
Эритробласты (нормобласты). На дальнейших стадиях дифференцировки происходит
уменьшение размеров клетки, элиминация ядра, конденсация хроматина, потеря
органелл и РНК, постепенное увеличение содержания гемоглобина. Последовательно
различают эритробласты базофильные,
полихроматофильные и оксифильные.
Базофильный эритробласт мельче проэритробласта, его
диаметр колеблется в пределах 13-16 мкм. Ядро характеризуется наличием плотного
хроматина. Цитоплазма базофильна. Около ядра
находится клеточный центр. Сохраняется способность к митозу. Активно
синтезирует гемоглобин.
Полихроматофильный эритробласт имеет диаметр 12-15 мкм, содержит значительное
количество гемоглобина. Клетки продолжают синтезировать гемоглобин и сохраняют
способность к делению. Размеры ядра уменьшаются.
Оксифильный эритробласт имеет небольшие размеры(8-10 мкм) и ацидофильную
цитоплазму. Ядро небольшое, хроматин конденсированный. На ранних стадиях
нормобласты сохраняют способность к делению, но при переходе к стадии эритроидной клетки эта способность утрачивается. Наблюдается
вытеснение пикнотического (дегенерирующего) ядра.
Полностью разрушается белоксинтезирующая система.
Эритробластный островок.
В костном мозге выделяют отдельные структурные единицы эритропоэза
– эритробластные островки. Островок обычно состоит из
одного макрофага, окруженного клетками-предшественниками эритропоэза.
Макрофаг образует отростки, на поверхности которых расположены делящиеся эритроидные клетки. По мере дифференцировки эритроидная клетка мигрирует к концу отростка макрофага, а
следом за ней перемещаются менее дифференцированные клетки. Ядро выталкивается
и фагоцитируется макрофагами. Безъядерная клетка - ретикулоцит вступает
в контакт с эпителием ближайшего синусоида, проходит
через его стенку и попадает в общий кровоток.
Ретикулоциты содержат остатки рибосом и РНК. Они выходят в
кровоток и составляют до 1% общего числа циркулирующих эритроцитов. После
выхода в кровоток в течение первых 24-48 часов ретикулоцит
завершает созревание и превращается в эритроцит. При этом клетка приобретает форму
двояковогнутого диска, а последние сохранившиеся органеллы разрушаются
ферментами.
Процесс эритропоэза можно представить в виде следующей схемы:: Эритропоэтинчувствительная клетка → проэритробласт
→ базофильный эритробласт
→ полихроматофильный эритробласт
→ оксифильный эритробласт
(нормобласт) → ретикулоцит → эритроцит,
макрофаг.
Гранулоцитопоэз. Гранулоциты образуются в красном костном мозге.
Нейтрофилы и базофилы развиваются из полипотентной колониеобразующей единицы
нейтрофилов и базофилов; эозинофилы – из униполярной колониеобразующей единицы
эозинофилов. По мере дифференцировки размеры клеток уменьшаются, хроматин
конденсируется, изменяется форма ядра, в цитоплазме накапливаются гранулы.
В развитии
гранулоцита можно выделить 6 морфологически различных стадий: миелобласт, промиелоцит,
миелоцит, метамиелоцит, палочкоядерный
и сегментоядерный гранулоциты. Специфические гранулы появляются на стадии
миелоцита. С этого момента клетки
называют в соответствии с типом образующихся из них зрелых гранулоцитов.
Клеточные деления прекращаются на стадии метамиелоцита.
Миелобласт – малодифференцированная клетка диаметром около 15
мкм. Крупное округлое или овальное ядро содержит 1-3 ядрышка. Цитоплазма слабо базофильна, гранулы отсутствуют.
Промиелоцит формируется из миелобласта.
Крупные клетки диаметром 15-24 мкм. Округлое ядро располагается эксцентрично,
хроматин конденсирован. Цитоплазма базофильна и
содержит азурофильные гранулы.
Миелоцит. Размеры клеток меньше предыдущих (12-16 мкм).
Появляются специфические гранулы, что позволяет различать 3 типа миелоцитов – нейтрофильный, эозинофильный и базофильный. Образование и накопление гранул продолжается в
течение последующих клеточных делений. Ядро приобретает бобовидную форму,
хроматин более конденсирован.
Метамиелоцит. В результате деления миелоцитов образуются нейтрофильные, эозинофильные и базофильные
метамиелоциты. Размеры этих клеток составляют 12-14
мкм. Содержание специфических гранул больше, чем на предыдущей стадии. В ядре
появляются глубокие вырезки. Хроматин сильно конденсирован. Утрачивается
способность к митозу.
Палочкоядерный гранулоцит.
Метамиелоциты дифференцируются в палочковидные
гранулоциты – клетки непосредственно предшествующие зрелым формам. Их размер
составляет 10-12 мкм. Ядро имеет подковообразную форму. Они могут выходить в
кровоток и составляют 3-5% общего количества циркулирующих лейкоцитов.
Сегментоядерный гранулоцит. Палочкоядерные гранулоциты
дифференцируются в зрелые формы – сегментоядерные. Сегментированное ядро
содержит плотный хроматин.
Схема гранулоцитопоэза:
а) миелобласт
→ промиелоцит → нейтрофильный
миелоцит → метамиелоцит → палочкояддерный нейтрофил → сегментоядерный
нейтрофил.
б) миелобласт → промиелоцит
→ эозинофильный миелоцит→ метамиелоцит→
палочкояддерный эозинофил→ сегментоядерный
эозинофил
в) миелобласт → промиелоцит → базофильный
миелоцит → метамиелоцит → палочкоядерный базофил → сегментоядерный базофил
Моноцитопоэз. Моноциты и гранулоциты имеют общую клетку –
предшественницу - колониеобразующую единицу гранулоцитов и моноцитов,
образующуюся из полипотентной колониеобразующей единицы миелопоэза.
При развитии моноцитов выделяют две стадии – монобласт
и промоноцит. До достижения стадии зрелого моноцита
клетки проходят 3 деления. Постепенно уменьшается размер клеток, появляется
углубление в ядре. Все зрелые моноциты покидают костный мозг. В течение 2-4
суток они находятся в кровотоке, а затем мигрируют в ткани. Моноциты могут
дифференцироваться в клетки моноцитарно-макрофагической
системы (макрофаги, микроглиоциты, остеоциты).
Лимфоцитопоэз. Из стволовой кроветворной клетки образуется
полипотентная колониеобразующая единица лимфоцитопоэза.
В-лимфоциты образуются в костном мозге. Созревание Т-лимфоцитов происходит в
тимусе. При образовании лимфоцитов выделяют две стадии: лимфобласт
и пролимфоцит. Лимфобласт
крупнее зрелого лимфоцита.
Главная особенность лимфоцитопоэза
- постоянное уменьшение объема клетки. В
отличие от других клеток крови лимфоциты могут пролиферировать и вне костного
мозга. Это происходит в тканях иммунной системы в ответ на антигенную
стимуляцию.
Дифференцировка В–лимфоцитов. Выделяют 6 стадий созревания В-лимфоцитов:
про-В-клетка, пре-В-клетка, экспрессирующая
мембранные иммуноглобулины, активированная В-клетка, В-лимфобласт,
плазматическая клетка, секретирующая иммуноглобулин.
Дифференцировка Т-лимфоцитов оооисходит в тимусе из
клеток-предшественниц, поступающих в вилочковую железу из костного мозга.
Зрелые Т-лимфоциты покидают тимус. Они поступают в периферическую кровь и
лимфоидные органы.
Тромбоцитопоэз. Тромбоцитам дают начало самые крупные клетки
костного мозга - мегакариоциты, имеющие размеры 30-100 мкм. Тромбоциты
формируются путем фрагментации цитоплазмы мегакариоцитов.
Мегакариобласт – предшественник мегакариоцита, образующаяся
из полипотентной колониеобразующей единицы миелопоэза.
Мегакариоцит образуется из мегакариобласта. Для него
характерно полиплоидное дольчатое ядро с диффузно распределенным хроматином.
Цитоплазма слабо базофильна и содержит мелкую
зернистость. В цитоплазме зрелого мегакариоцита различают 3 зоны: перинуклеарную, промежуточную и наружную. Перинуклеарная зона содержит комплекс Гольджи,
гладкую и гранулярную ЭПС, гранулы, центриоли и трубочки. Эта зона остается
связанной с ядром после отделения тромбоцитов. Промежуточная зона содержит
развитую систему взаимосвязанных пузырьков и трубочек (демаркационная
мембранная система), переходящую в клеточную мембрану и выполняющую
разграничительную функцию при формировании тромбоцитарных
полей. Наружная (краевая) зона содержит элементы цитоскелета
и пересекается мембранами, связанными с системой демаркационных мембран.
Факторы гемопоэза. Образование клеток крови стимулирует гемопоэтические
факторы роста – фактор стволовых клеток, колониестимулизирующие
факторы, интерлейкины, эритропоэтин,
лептин, тромбопоэтин. На кроветворение влияют
фолиевая кислота и витамин В12, участвующие в синтезе ДНК в ходе
массового образования новых клеток крови. Дифференцировку кроветворных клеток
контролируют факторы транскрипции.
Теории кроветворения
Существует несколько теорий о
происхождении форменных элементов крови: унитарная, дуалистическая, триалистическая, полифилетическая.
Согласно унитарной теории (А.А. Максимов, Ф. Вейденрейх, А.Феррата), все
форменные элементы крови происходят от одной родоначальной
клетки - гемоцитобласта. Их разнообразие объясняется условиями обмена веществ в
каждом органе кроветворения, что вызывает стимуляцию образования одних и
подавление других клеточных форм.
Согласно дуалистической теории (О.
Негели, Г.Шридде, А. Поппангейм, А.А. Заварзин),
форменные элементы крови могут возникать из двух родоначальных
клеток: из лимфобластов - лимфоциты и моноциты, и из миелобластов – эритроциты и гранулоциты.
По мнению триалистической
теории (Л.Ашофф, К. Кийоно),
все кровяные клетки происходят из трех родоначальных
клеток: лимфоциты образуются из лимфобластов,
моноциты - из ретикулоэндотелия, гранулоциты и
эритроциты – из миелобластов.
Наиболее признанной является унитарная теория.
В настоящее время общепризнано, что родоначальной
клеткой всех форменных элементов крови является полипотентная стволовая
кроветворная клетка – гемоцитобласт, которая дает начало клеткам -
предшественницам для всех форм гемопоэза.
Клетки–предшественницы связаны гемопоэтическими факторами питания и
нейрогуморальной регуляцией, а при изменении наблюдается нарушение гемопоэза.
Эндотелий выстилает
внутреннюю поверхность сердца, кровеносных и лимфатических сосудов. Он состоит
из плоских клеток, расположенных в один ряд, и напоминает строение однослойного
однорядного плоского эпителия (мезотелия). Границы между эпителиальными
клетками имеют вид извилистых линий. Клетка имеет одно или несколько ядер,
митохондрии, ЭПС, хорошо развит комплекс Гольджи.
Эндотелиальные клетки расположены на базальной мембране, которая отделяет их от
подлежащей соединительной ткани. Базальная мембрана обеспечивает обмен веществ с кровью (лимфой) и другими элементами стенки сосуда.
Эндотелий образуется из мезенхимы, которая
способна к митотическому делению. В
кроветворных органах и печени, где ток крови замедляется
и кровеносные капилляры расширяются, превращаясь в синусоиды и синусы, строение
и функция эндотелия отличается от таковых в других отделах сосудистой системы.
В синусах и синусоидах границы между эндотелием и ретикулярными клетками
выражены слабо в связи с отсутствием в ряде мест базальной мембрны.
Здесь эндотелиальные клетки способны к фагоцитозу, в отличие иных частей
сосудистой системы.
При повреждении кровеносного или лимфатического
сосуда, которое не сопровождается разрушением и гибелью эндотелиальных клеток,
они делятся и возникают многоядерные клетки. При гибели эндотелиальных клеток
они отторгаются, а соседние клетки растягиваются и заполняют дефект. В случае
возникновения значительного дефекта он быстро ликвидируется за счет размножения
соседних эндотелиальных клеток.
Ретикулярная ткань (reticulum –
сетка) имеет много общих черт с мезенхимой и прежде всего сетевидное строение.
Ретикулярная ткань состоит из основного вещества, ретикулиновых волокон и
ретикулярных клеток, имеющих длинные отростки. Ретикулярные клетки вместе с
ретикулиновыми волокнами образуют рыхлую сеть.
Многочисленные отростки ретикулярных
клеток контактируют с другими ретикулярными клетками, переплетаются с ними,
обусловливая сетевидное строение ткани. Ретикулярные клетки – крупные клетки с оксифильной цитоплазмой, ядром округлой формы с одним или
несколькими ядрышками. Клетки имеют развитую гранулярную ЭПС, аппарат Гольджи, цитоскелет. Ретикулоциты синтезируют коллаген типа ІІІ,
гемопоэтические факторы роста,
регулирующие деление и дифференцировку кровяных клеток. Они выполняют и
трофическую функцию - распределение питательных веществ, поступающих из
кровеносных капилляров. Ретикулярные волокна состоят из коллагена типа ІІІ.
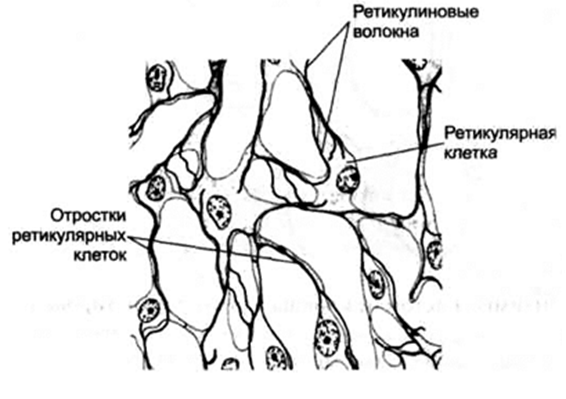
Рис. 17. Ретикулярная ткань
Ретикулярная ткань встречается в органах
кроветворения и иммуногенеза, образуя их строму. Она создает специфическое
микроокружение для развивающихся клеток крови. Ретикулярные клетки способны к
фагоцитозу. При старении организма количество ретикулярной ткани уменьшается,
т.к. она превращается в другие виды соединительной ткани.
Ретикулярная ткань развивается из
мезенхимы. В процессе эмбрионального гистогенеза клетки мезенхимы зародыша
дифференцируются в ретикулобласты, способные
образовывать ретикулиновые волокна – первые опорные элементы, которые
обеспечивают функционирование тканей зародыша. Впоследствии ретикулобласты
превращаются в ретикулоциты. Последние
могут дифференцироваться в макрофаги, приобретая способность к фагоцитозу.
Наряду с клетками мезенхимы, ретикулобласты и ретикулоциты дают начало клеткам малодифференцированного
эндотелия - ретикулоэндотелию, т.е. ретикулярным
клеткам, которые наряду с клетками эндотелия образуют выстилку в венозных и
лимфатических синусах. Клетки ретикулоэндотелия
дифференцируются в перициты кровеносных и лимфатических сосудов, венозных и
лимфатических синусов. Перициты в течение онтогенеза сохраняют потенции
дифференциации в ретикулярые клетки, являясь основным
источником для регенерации, а также для восстановления других клеточных форм
соединительной ткани.
Мезенхима – эмбриональная соединительная ткань - источник
происхождения клеток всех соединительных тканей. Мезенхимные клетки имеют
звездчатую или веретеновидную форму с нежними
ветвящимися отростками, формирующими сеть. В овальном ядре хроматин
диспергирован и имеются хорошо заметные ядрышки. В клетке много рибосом и
полисом. Гелеобразный внеклеточный матрикс состоит из основного вещества и
небольшого количества рассеянных волокон (в основном ретикулиновых). В
определенных условиях мезенхимные клетки могут отделяться друг от друга, приобретать
способность к амебоидному движению и фагоцитозу.