Глава 4.
Этническая антропология (расоведение)
4.1. Расы и их классификация
4.2. Расообразование, причины и факторы
4.3. Смешение,
адаптация и изоляция как факторы формирования рас
Контрольные
вопросы
4.1. Расы и их классификация
Расоведение
изучает
антропологический состав народов земного шара в настоящем и прошлом. Полученные
в результате этого материалы дают возможность выяснить родственные отношения
между расами и историю их возникновения, место и причины происхождения расовых
типов, их расселения по планете.
С биологической точки зрения человечество Земли представляет собой
один вид Homo sapiens, который распадается на ряд более мелких делений,
называемых расами.
Вид (species)
– основная
таксономическая категория в биологической систематике, рассматривается как
совокупность особей, почти всегда состоящая из большего или меньшего числа
подвидов или географических больших рас, занимающих определенную территорию,
или ареал. Так, в частности, по строению черепа отмечены расы горилл, расы
белого медведя, расы дикого северного оленя, бобра и т. д. Признаки, по
которым различаются расы, обычно сравнительно устойчивы, т. е. мало меняются
под непосредственным воздействием среды.
Хотя
принципы выделения рас у человека и животных представляют некоторую очевидную
аналогию (в наличии ареала, наборе расовых таксономических признаков, в их относительной
устойчивости, независимости варьирования), однако их значение и история
принципиально иные у человека и животных. Процесс расообразования у человека
протекал под влиянием двух факторов: биологического и социального.
Механизмы
образования отдельного расового признака относятся у человека к области
биологии. Конкретная же история формирования расовых типов относится к области
социальной жизни человека. Так, например, для понимания механизма расщепления
признаков человека, определяющих цвет и форму волос, достаточно знания
некоторых общих и частных положений генетики. Для понимания причин проникновения,
например, тугой формы волос за пределы Восточной Азии на остров Мадагаскар,
необходимо знать историю заселения этих областей, а следовательно, те социальные
причины, которые его вызывали.
Основной
особенностью расовых вариаций, в отличие от всех других внутривидовых вариаций,
является географический ареал их распространения. В пределах этого ареала
сосредоточена обычно большая часть представителей данного расового типа.
Необходимо
строго различать два весьма разных понятия – нация и раса. Людей объединяет в нации
общность языка, территории, экономической жизни, психического уклада. В
отличие от нации, раса есть
совокупность популяций, обладающих общностью физического типа, происхождение которого
связано с определенным ареалом.
Слово раса происходит от арабского корня рас – голова, начало или итальянского
слова razza – племя. Впервые в современном
смысле слово раса встречается у
французского ученого Ф. Бернье (1684
г.).
В
настоящее время существует большое разнообразие в определении понятия «раса».
Приведем некоторые из них.
Раса – это группа людей, имеющая
общие физические черты.
Раса – это
совокупность людей, обладающих общностью физического типа, происхождение
которого связано с определенным ареалом (Рогинский, Левин, 1978).
Раса – это
«популяция» (или группа популяций), отличающаяся от других таких же популяций того
же вида частотами генов, хромосомными перестройками или наследуемыми
фенотипическими признаками (Айала, 1984).
Раса – это
большая популяция индивидуумов, у которых значительная часть генов общая и
которую можно отличить от других рас по общему для нее генофонду (Фогель, Мотульски,
1990).
Раса
человека – это
исторически сложившиеся группы людей, связанные единством происхождения, которое
выражается в общих наследственных морфологических и физиологических признаках,
варьирующих в определенных пределах. Так как групповая и индивидуальная
изменчивость этих признаков не совпадает, расы являются не совокупностями
особей, а совокупностями популяций, т. е. территориальных групп людей,
объединенных брачными связями.
Разнообразие определений находит отражение в многообразии расовых
классификаций. Во многом это связано с принципами, которые закладывают
исследователи в план построения своих классификаций.
Между тем общее в понятии «раса» – это
определенность ареала и физическое сходство.
Исторические источники донесли до нас факты того, что люди уже
достаточно давно обратили внимание на существование территориальных различий во
внешнем облике себе подобных. Традиционно приводится пример египтян, которые в
середине II тысячелетия до н. э. использовали четыре цвета при изображении
людей разного происхождения: красный – для
египтян, желтый – для
народов Востока, белый – для народов
Севера, черный – для
южных народов.
Для многих народов, как в древности, так и в наше время, основным
отличительным морфологическим признаком является цвет кожи. В антропологии к
существенным расовым признакам относятся форма волос головы, степень развития
третичного волосяного покрова (усы, борода), пигментация волос и кожи.
Несколько меньшее значение как расовым признакам придается росту,
весу и пропорциям тела. Немалое значение для выделения расовых типов имеет
изучение признаков головы и лица, формы носа.
Для антропологических исследований привлекаются и многие
специальные признаки: данные о генах, группах крови, флексорных линиях ладоней,
узорах на пальцах (папиллярные узоры), форме зубов и др.
Первая научная классификация рас принадлежит Франсуа Бернье (1684 г.), который в конце
XVII века разделил человечество на четыре расы (по современной терминологии – европеоиды, монголоиды, негроиды и лапоноиды, или индейцы Америки).
Карл Линней также выделил четыре расы человека (африканская, азиатская, американская и
европейская).
Первые три совпадают с классификацией Ф. Бернье, но четвертой у
него была не лапоноидная, а американоидная раса (индейцы). Бюффон добавил к ним
южноазиатскую.
В 1775 году Блуменбах выделил 5 наиболее известных «цветных» рас
человека:
- белую, или кавказскую (европеоидную);
- желтую, или монголоидную;
- черную, или эфиопскую;
- красную, или американоидную;
- коричневую, или малайскую.
В 1902 году была опубликована классификация И. Деникера, которая
считается основополагающей для очень многих последующих. Он вводит понятие
«соматологических единиц», к которым относит отдельных особей. «Эти единицы, – пишет
Деникер, – служат теоретическими типами, состоящими из совокупности физических
признаков, сочетанных друг с другом определенным образом».
Эти типы он называл расами и подрасами. Деникер разделил
человечество на 29 рас, объединив их в шесть групп («большие расы», по
современной терминологии). Ведущими признаками были приняты форма волос и цвет
кожи.
Внутри групп расы выделены на основании цвета кожи, формы головы,
лица и носа, других морфологических признаков:
1) шерстовидные
волосы, широкий нос (расы – бушменская, негритосская,
негрская, меланезийская);
2) курчавые
или волнистые волосы (расы – эфиопская, австралийская,
дравиддийская, или мелано-индийская, ассироидная);
3) волнистые,
темные или черные волосы и темные глаза (расы – индо-афганская,
арабская, берберская, средиземно-приморская, островная-иберийская, западная,
адриатическая);
4) волнистые
или прямые волосы, блондины со светлыми глазами (расы – северная,
восточная);
5) прямые
или волнистые черные волосы, глаза темные (расы – айнская,
полинезийская, индонезийская, южноамериканская);
6) прямые
волосы (расы – североамериканская, среднеамериканская, патогонская, эскимосская,
лопарская, угорско-елисейская, туранская, монгольская).
Несмотря на некоторые несоответствия классификации Деникера
(например, ассирийцы попали в одну группу с папуасами), большинство его
«диагнозов» подтвердилось.
В нашей стране наиболее известны три классификации. Авторами этих
классификаций являются: В.В. Бунак (Наука о расах и расизм, 1939), Г.Ф. Дебец
(Современная этнография, 1958, № 4), Я.Я. Рогинский и М.Г. Левин (Антропология,
1978).
Классификация Я.Я. Рогинского и М.Г. Левина имеет довольно строгую
структуру: три большие расы (евразийская, азиатско-американская,
экваториальная) подразделяются на малые расы, промежутки между большими
заполнены шестью промежуточными расами. В названиях соблюден географический
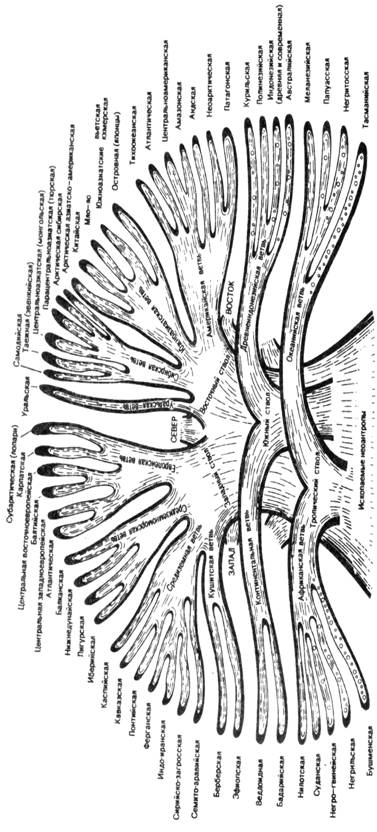
принцип. Символ схемы – круг (рис. 110), т. е.
во главу угла ставится географическое распределение вариации признаков, а не
индивидуальная типология.
По иерархическому принципу построены классификации
Г.Ф. Дебеца и В.В. Бунака, в которой несколько основных, «больших единиц»
последовательно подразделяются на более мелкие
категории (малые расы, антропологические типы и т. п.).
В логике построения они имеют тройную иерархию. Символ схемы
классификации Г.Ф. Дебеца – дерево (рис. 111), где переплетение и слияние
ветвей отражают процессы смешения при образовании антропологических типов.
Для классификации В.В. Бунака характерен отказ от принципа
дихотомии при ветвлении (различий) смежных групп. Символом схемы его классификации
служит куст (рис. 112).

Рис. 110. Схема расовой классификации (по Рогинскому, Левину, 1978). На
схеме видно, что промежуточное место между экваториальной и евразийской
большими расами занимают две расы: эфиопская и южноиндийская. Между евразийской
и азиатско-американской размещаются также две расы – южносибирская и уральская;
наконец, особое место по значительной части признаков, промежуточное между
экваториальной и азиатско-американской, занимают полинезийская и курильская
расы
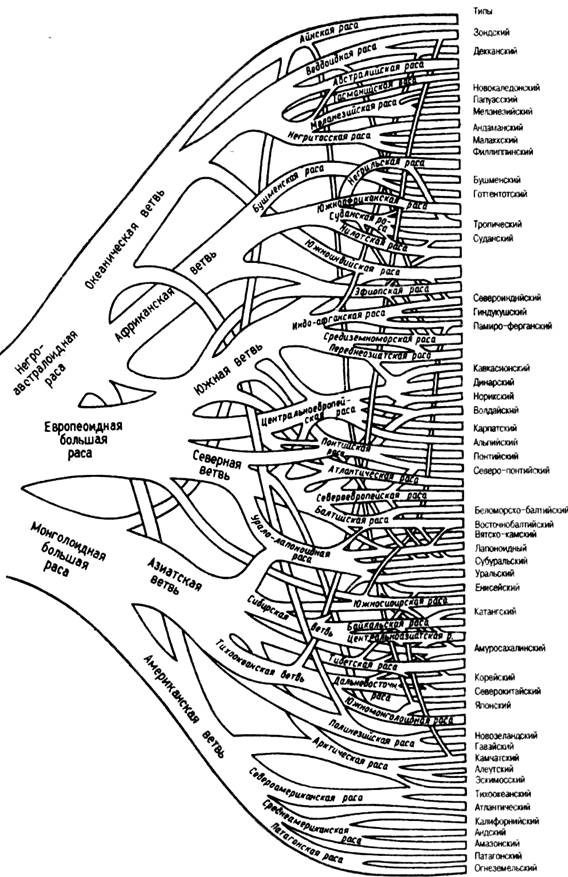
Рис. 111. Схема классификации человеческих рас по Г.Ф. Дебецу (1958 г.). Переплетение и
слияние ветвей отражают процессы смешения, имевшие место в различные периоды
эволюции современного вида
4.1.1.
Современная классификация рас
Несмотря
на большое количество разных классификаций между ними существует сходство по
многим антропологическим типам и их группировке в более крупные подразделения.
Практически во всех классификациях упоминаются следующие антропологические типы
независимо от их ранга:
- Европеоиды (еврозийская, кавказоидная раса), среди
которых обязательно выделяют светло- и темнопигментированные, а также лопарский
варианты;
- Монголоиды
(азиатско-американская) с американоидными эскимоидным и дальневосточным
типами;
- Негроиды
(экваториальная) с австралоидным, меланезийским, койсанским, каноидным и
пигмейским (негрилльским, центральноафриканским) типами.
В
самостоятельные варианты со спорным происхождением выделяются:
- полинезийская;
- айнская (курильская);
- эфиопская
(восточноафриканская);
- веддоидная;
- дравиддийская
(южноиндийская).
К
смешанным причисляются:
- южносибирская (туранская);
- уральская (угорская).
Часто в
категорию самостоятельных или смешанных рас попадают:
- австралийская;
- бушменская;
- лопарская;
- негритосская и негрилльская;
- американоидная.
Ниже
приводится морфологическое описание «больших» и «малых» рас.
I.
Европеоидная большая раса
Европеоидная
раса самая большая по численности (2/3 общей численности населения Земли) и
хорошо изученная.
В
настоящее время европеоиды населяют практически всю обитаемую сушу. До эпохи
Великих географических открытий европеоидные антропологические типы занимали
территории Европы, Северной Африки, Передней и Средней Азии, Среднего Востока и
Индии.
Границы
ареала европеоидной расы подвижны, а в «контактных» зонах, населенных другими
антропологическими типами, возникли формы смешанного происхождения.
Вся зона
заселения европеоидов разделена горными массивами в направлении с запада на
восток: Пиренеи – Альпы – Балканы – Кавказ – Копетдаг – Тянь-Шань.
На
антропологические особенности европеоидной расы оказали влияние умеренный и
средиземноморский климат и то обстоятельство, что северные районы Европы и некоторые
горные районы сравнительно недавно (10 тыс. лет назад) окончательно
освободились от ледника. Открылись новые площади для заселения и возможности
контакта южных и северных групп населения и их смешение. Формирование и современная
локализация европеоидной расы главным образом были обусловлены событиями,
происшедшими в неолите.
Европеоидная
раса подразделяется на ряд малых рас или типов (рис. 113).
1. Атланто-балтийская раса. Основной ареал – Скандинавия, Британские острова, северные
районы Западной и Восточной Европы. Для популяций основного ареала данной расы
характерна светлая пигментация кожи. Частота светлых глаз может достигать 75 %,
а светлых волос – 50 %.
Волосы
широковолнистые и прямые, мягкие, нос обычно прямой и узкий, с высоким
переносьем. Третичный волосяной покров на теле – от среднего до слабого. Головной указатель – на границе долихо- и
мезокефалии, рост высокий, высота лица несколько преобладает над шириной.
2. Беломоро-балтийская раса. Зона наибольшей частоты распространения – от Балтийского до Белого
моря. Раса светлопигментированная, особенно относительно цвета волос. Третичный
волосяной покров несколько слабее, чем у атланто-балтийской расы.

Рис. 113. Евразийская большая раса: 1 –
атланто-балтийская; 2 – среднеевропейская; 3 – индо-средиземноморская; 4 –
балкано-кавказская;
5 – беломорско-балтийская
Рост
средний. Лицо шире и ниже. Головной указатель выше (тенденция к брахикефалии)
Нос более короткий с прямой или вогнутой спинкой (до 30–40 % случаев) является
главным физиологическим отличием от атланто-балтийской расы.
3. Среднеевропейская раса. Ареал распространения этой расы –
Североевропейская равнина от Атлантики до Волги. По антропологическим признакам
близка к предыдущим, но характеризуется более темной пигментацией волос.
Большинство пропорций тела имеют средние значения. Головной указатель, как
правило, умеренно брахикефальный. Третичный волосяной покров на теле умеренный.
Нос с прямой спинкой и высоким переносьем, длина его варьирует.
Многие
исследователи полагают, что среднеевропейская раса имеет смешанное
происхождение.
4. Балкано-кавказская малая раса. Популяции данного антропологического типа
занимают в основном район Евразийского горного пояса.
Длина
тела средняя и выше средней. Волосы темные, чаще волнистые. Глаза темные и
смешанного типа. Частота встречаемости светлых глаз в разных группах – от 2 до 20 %. Третичный
волосяной покров развит сильно (в некоторых популяциях мировой максимум).
Головной указатель брахикефального типа. Пропорции лица варьируют, ширина лица – от средней до выше средней.
Нос крупный, часто с выпуклой спинкой.
5. Индо-средиземноморская малая раса. Индо-средиземноморская малая
раса распространена в некоторых южных районах Европы, Северной Африке, Аравии и
других южных районах от Евразии до Индии. Цвет кожи смуглый.
Волосы
темные, волнистые. Глаза темные и смешанные (до 10–25 %), светлые очень редки.
Третичный волосяной покров развит умеренно. Нос прямой и узкий с высоким переносьем.
Лицо узкое. Головной указатель долихо- и мезокефалийного типа.
II.
Монголоидная
(азиатско-американская) большая раса
Ареал
расселения монголоидных популяций охватывает Восточную Азию, Индонезию,
Центральную Азию, Сибирь, Америку. Эта раса отличается смуглыми или светлыми оттенками
кожи. Цвет кожи становится более темным по мере продвижения от полюсов к
экватору. Пигментация волос варьирует в пределах темных оттенков шкалы. Волосы
прямые, нередко жесткие. Рост бороды и усов слабый. Ширина носа средняя с
низким или средним по высоте переносьем. Азиатские монголоиды характеризуются
слабо выступающим носом, в отличие от американских. Лицо уплощенное с сильно
выступающими скулами, размеры лица крупные. Частота эпикантуса уменьшается при
продвижении к Юго-Востоку Азии и отсутствует у американских монголоидов.
Азиатско-американская раса подразделяется на несколько малых рас (рис. 114).
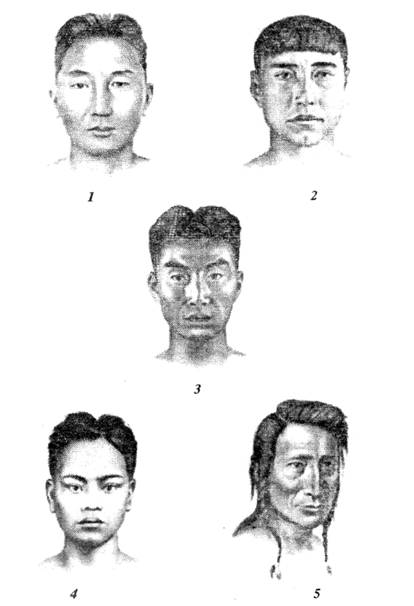
Рис. 114. Азиатско-американская раса: 1 – североазиатская; 2 – арктическая
(эскимосская); 3 – дальневосточная; 4 – южноазиатская; 5 – американская
1. Североазиатская малая раса. Западная граница ареала проходит по Енисею,
южная граница примыкает к центральноазиатским пустыням и горам Хингана и Станового
хребта, северные и восточные границы океанические. Черты североазиатской расы
лежат в основе антропологического типа многих коренных народов Сибири: эвенков,
якутов, бурятов и т. д. Цвет кожи более светлый, чем у других представителей
монголоидной расы. Волосы темные и темно-русые, обычно прямые и жесткие. У
эвенков волосы мягкие. Цвет глаз от темного до светло-карих оттенков. Лицо, как
правило, высокое и широкое, очень плоское. Мозговой череп с большим
горизонтальным диаметром и малым высотным. Нос варьирует по величине и степени
выступания. Часто встречается эпикантус.
Длина тела средняя и ниже средней. В составе североазиатской расы выделяются
два морфотипа – байкальский и центральноазиатский.
Байкальский тип характеризуется максимальной
монголоидностью в морфологии лица (максимальная уплощенность) и минимальной
пигментацией, слабым ростом бороды, низким переносьем и тонкими губами.
Центральноазиатский тип представлен в различных вариантах,
некоторые из которых сближаются с байкальским типом, другие – с вариантом арктической и
дальневосточной рас.
2. Арктическая малая раса. Ареал распространения – от Верхоянского хребта на
западе до Гренландии. Южная граница проходит примерно по 60° северной широты.
Этот тип характеризуется более темной пигментацией, чем у североазиатской расы,
прямыми жесткими волосами и наличием эпикантуса (от 30 до 50 %). Лицо имеет
пентагоноидное очертание с более выступающим носом, толстыми губами, меньшей
скуловой шириной.
Головной
указатель долихокефального типа. К этой малой расе принадлежат эскимосы, чукчи,
американские индейцы, коряки.
3. Дальневосточная малая раса. В значительном проценте – это жители Китая, Кореи,
Японии. Кожа смуглая, глаза темные, волосы густые и прямые, жесткие и очень
темные. Эпикантус встречается от 70 до 95 % случаев. Третичный волосяной покров
развит слабо. Длина тела средняя или выше средней. Лицо узкое, нос довольно
длинный с прямой спинкой, слабо- или средневыступающий.
4. Южноазиатская малая раса. Распространена в странах Южной и Юго-Восточной
Азии. Цвет кожи более темный, чем у дальневосточной расы. Губы толстые, характеризуется
меньшим ростом, более высоким носовым указателем, менее уплощенным лицом, чем у
дальневосточной расы. Эпикантус встречается реже (20–50 %). Мозговой череп
небольшой и относительно широкий.
5. Американская раса. Занимает обширную территорию Американского
континента с самыми различными экологическими условиями. Несмотря на то, что
внутри расы выделяют несколько морфотипов, она достаточно едина в основных
чертах.
Для
большинства популяций и морфотипов американских индейцев характерен крупный
нос, иногда с выпуклой спинкой. Эпикантус почти отсутствует, кожа очень смуглая.
Размеры лица крупные с меньшей уплощенностью. Длина тела средняя и большая.
Нетипичность
американских индейцев дает некоторым исследователям основание выделять ее в
отдельную большую расу.
III.
Австрало-негроидная
(экваториальная)
большая раса
До эпохи
европейской колонизации экваториальная большая раса распространялась главным
образом к югу от тропика Рака в Африке, а также включала некоторые популяции
Индонезии, Новой Гвинеи, Меланезии и все аборигенное население Австралии.
Ведущим признаком считается темный цвет кожи. Волосы волнистые или курчавые.
Нос маловыступающий, широкий с низким или средним переносьем. Губы толстые, ротовая
щель большая.
Большая
экваториальная раса делится на ряд малых рас (рис. 115).
1. Негрская малая раса. Популяции этой расы наиболее многочисленны в
Африке. Цвет кожи темный и очень темный. Глаза темные. Волосы сильно курчавые и
спирально завитые. Нос широкий в крыльях, с низким и плоским переносьем. Губы
толстые, вывернутые. Третичный волосяной покров средний и слабый. Глазная щель
широко открыта, глазное яблоко несколько выступает вперед. Длина тела средняя и
выше средней.

Рис. 115. Экваториальная большая раса: 1 –
австралийская; 2 – веддоидная; 3 – меланезийская; 4 – негрская; 5 –
негрилльская (центральноафриканская); 6 – бушменская (южноафриканская)
2. Бушменская (южноафриканская) малая раса. Обитает в пустынных и полупустынных районах Южной Африки. Отличается от
негрской расы более светлой кожей и низким ростом. Цвет кожи желтовато-бурый.
Волосы спирально завитые. Третичный волосяной покров слабый. Нос широкий с
низким переносьем. Разрез глаз меньше, чем у негрской расы, встречается
эпикантус. Лицо небольшое уплощенное. Длина тела ниже средней. Сильный
поясничный лордоз сочетается со стеатопигией
(отложением жира в ягодичной области).
3. Негрилльская (центральноафриканская) малая раса. К ней принадлежат популяции
аборигенного населения тропического леса Африки. Пигментация и форма волос как
у негрской расы. В отличие от негрской
расы, характеризуется низким ростом, более обильным развитием третичного
волосяного покрова, тонкими зубами, резко выступающим широким носом.
4. Австралийская малая раса. Представлена аборигенами Австралии. В отличие
от негрской расы, цвет кожи не столь темный, цвет волос варьирует от коричневых
до черных. У детей и молодых женщин (до 20 %) встречается светловолосость.
Форма волос широковолнистая, узковолнистая и локоновая. Глаза сильно
пигментированные. Разрез глаз большой, но глазное яблоко посажено очень
глубоко. На лице хорошо развит третичный волосяной покров, на теле – умеренно. Нос широкий в крыльях с низким
переносьем. Губы средней толщины. Есть прогнатизм. Рост средний и высокий.
5. Меланезийская малая раса. Распространена на Новой Гвинее и островах
Меланезии. Отмечается политипия внутри расы. Характерной особенностью является
преобладание курчавых волос, в отличие от австралийской расы. По обильному
развитию третичного волосяного покрова и сильно выступающим надбровным дугам
некоторые ее варианты весьма сходны с австралийской расой. Обладают маленьким
ростом, встречаются очень малорослые группы (негрито).
6. Веддоидная малая раса. Распространена на островах Индонезии, в
Шри-Ланка и Южной Индии. Представляет собой уменьшенный (грацильный) морфотип
австралоидов. Кожа умеренно темная, волосы волнистые, толщина губ средняя,
умеренный прогнатизм. Третичный волосяной покров довольно слабый. Длина тела
средняя и ниже средней.
Пограничными
между евразийской и экваториальной расами считаются эфиопская и южноиндийская.
Эфиопская
(восточноафриканская) раса. Цвет кожи варьирует от светло-коричневого до
темно-шоколадного. Волосы чаще курчавые,
но менее спирально завитые, чем у негров.
Третичный
волосяной покров ослаблен. Нос прямой широкий, с довольно высоким переносьем.
Рост бороды слабый или средний. Лицо узкое, губы умеренно толстые. По чертам
лица эта раса гораздо ближе к евразийской.
Предполагают,
что это древний своеобразный вариант, который в последующие эпохи постоянно
испытывал влияние потока генов от европеоидов и экваториалов.

Рис. 116. Переходные расы: 1 – эфиопская (восточноафриканская);
2 – южноиндийская
(дравиддийская); 3 – уральская; 4 – южносибирская (туранская); 5 –
полинезийская; 6 – курильская (айнская)
Южноиндийская (дравиддийская)
малая раса. Локализована
в Южной Индии на стыке южных европеоидов и веддоидов.
Кожа коричневых оттенков. Глаза и волосы темные. Волосы прямые и
волнистые. Лицо немного ниже и чуть-чуть шире, чем у эфиопской расы.
О смешанном происхождении эфиопской и южноиндийской малых рас
говорят данные частот по группам крови и сывороточным белкам.
В зоне контакта европеоидов и монголоидов выделяют две малые расы – уральскую и южносибирскую (рис. 116).
Уральская
малая раса. Ареал обитания – Приуралье, Зауралье и
частично Западная Сибирь. Уральская раса составляет основу антропологического
типа манси, хантов, селькутов, некоторых поволжских народов, а также народов
Алтае-Саянского нагорья. Характеризуется уральская раса преимущественно светлым
цветом кожи. Волосы темные и темнорусые, прямые и широковолнистые, часто
мягкие. Цвет глаз преимущественно смешанных и темных оттенков, но есть и
небольшой процент светлых.
Эпикантус встречается в 10–20 % случаев.
Нос прямой или с вогнутой спинкой, переносье средней высоты. Лицо небольшое и
относительно широкое, умеренно уплощенное. Губы средней толщины. Третичный
волосяной покров ослаблен. Рост средний и ниже среднего. По многим признакам
уральская раса занимает среднее положение между беломорско-балтийской и
североазиатской расами. По мнению некоторых исследователей, уральская раса,
возникла в результате смешения европеоидной и монголоидной групп в древности
(мезолит-неолит).
Южносибирская
(туранская) малая раса. Локализована в степях Казахстана, горных
районах Тянь-Шаня и Алтая-Саянах. Цвет кожи смуглый и светлый. Пигментация
волос и глаз немного темнее по сравнению с уральской расой. При нерезкой
выраженности монгольских черт в этой расе наблюдаются очень крупные размеры как
высоты лица, так и скуловой ширины, не меньше, чем у некоторых вариантов
североазиатской расы. Волосы чаще прямые и жесткие. Нос прямой, крупный,
переносье средней высоты.
Южносибирская
раса имеет смешанное происхождение. Начало смешения европеоидов и монголоидов в
евразийских степях достаточно точно датируется серединой 1 тыс. до н. э.
По всем
основным морфологическим, физиологическим и психическим особенностям,
характерным для современных людей, сходство между всеми расами велико, а различия
несущественны.
Лишены
всякого фактического основания концепции о существовании «высших» и «низших»
рас и об их происхождении от разных корней. Данные антропологии и других наук
доказывают, что расы происходят от одного вида ископаемых гоминид.
Неограниченные
возможности смешения (метисации) и социально-культурная полноценность смешанных
групп служат вескими доказательствами
видового единства человека.
Предполагают,
что современные расы начали складываться в позднем палеолите, и с тех пор они
все время изменяются вследствие кровосмешений и других причин.
4.2. Расообразование, причины и факторы
Признанной
в настоящее время гипотезой прародины человека, основанной на данных
антропологии и других наук, является гипотеза широкого моноцентризма (Рогинский,
1949).
Данная
гипотеза основана на признании того, что все расы происходят от одного вида
ископаемых гоминид, общности социально-психического, одинакового уровня физического
и умственного развития всех рас.
Предполагается,
что формирование рас произошло на территориях Восточного Средиземноморья,
Передней и, возможно, Южной Азии в эпохи раннего или среднего палеолита (около
200 тыс. лет назад), т. е. совпадает с возникновением человека современного
типа. Следовательно, основные расовые сочетания в западных и восточных районах
Старого Света складывались одновременно с оформлением признаков, присущих
современному человеку, а также с переселением части человечества в Новый Свет.
На
втором этапе происходило выделение вторичных очагов расообразования и формирование
ветвей в пределах основных расовых стволов в эпохи верхнего палеолита и частично
мезолита (около 15–20 тыс. лет назад).
На
третьем этапе расообразования происходило становление локальных рас. По времени
это соответствует началу мезолита и неолита (около 10–12 тыс. лет назад).
На
четвертом этапе возникли очаги расообразования и сформировались популяции с
углубленной расовой дифференциацией, сходной с современной. Это началось в
эпоху бронзы и раннего железа, т. е. в IV–III тыс. до н. э.
Каждый
представитель Homo sapiens sapiens обладает набором видоспецифических
признаков, отличающих его от представителей других видов, – это способность к прямохождению,
отсутствие волосяного покрова на большей части тела, высокоразвитый интеллект и
способность к речевой коммуникации. Помимо видоспецифических особенностей каждый
индивид несет целый набор индивидуальных, присущих только ему качеств, характеристик
– цвет волос и глаз, рост и телосложение, особенности поведения.
По
выражению американского ученого Роберта Пломина каждый человек есть уникальный
генетический эксперимент, который никогда больше не повторится.
Ученые
подсчитали, что число возможных сочетаний генов человека составляет 3 ×
1047, тогда как число живших на Земле людей за всю историю
человечества составляет
7 × 1010 , что на десятки порядков меньше.
Поэтому
на земном шаре практически не было и не может быть двух людей с одинаковым
набором генов. Вероятность того, что дети одних и тех же родителей (за исключением
монозиготных близнецов) будут иметь одинаковый набор генов, равняется одному
шансу на 64 триллиона возможностей.
В
зависимости от типа изменчивости в популяциях можно выделить признаки двоякого
рода:
- качественные (или дискретные),
характеризующиеся прерывистым рядом изменчивости;
- количественные (или непрерывные),
характеризующиеся континуальностью проявлений.
К
качественным признакам относятся, например, группы крови, а также признаки
гемофилии, альбинизма, определяющие грубую патологию, сенсорная
чувствительность (чувствительность к вкусу фенилтиомочевины и др.). Различия
между индивидуумами по дискретным признакам четко выражены, носители разных
вариантов качественных признаков в популяции могут встречаться с разной
частотой (рис. 117).
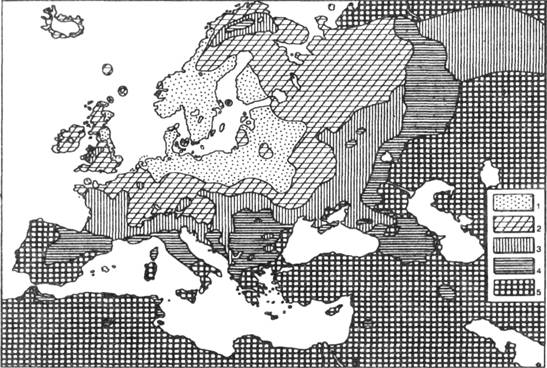
Рис. 117. Географическое распространение пигментации волос и глаз в Европе:
1 – существенное преобладание светлых и смешанных оттенков; 2 – светлых и
смешанных оттенков больше, чем темных;
3 – одинаковая частота светлых со смешанными и темными; 4 – темных больше, чем
светлых и смешанных; 5 – темные преобладают (Coon, 1939)
Количественная
изменчивость проявляется в таких морфологических признаках, как рост, вес, цвет
волос и кожи, и в таких поведенческих признаках, как интеллект, черты
темперамента и т. д.
Частотное
распределение по количественному признаку, проявляющему непрерывную
изменчивость, примерно соответствует кривой нормального распределения (см.
раздел морфологии).
Большинство
людей в популяции попадают в среднюю часть распределения, а на его краях
оказывается малая часть.
Индивидуальная
изменчивость на генотипическом и фенотипическом уровнях представляет собой
важную предпосылку эволюции, но это не сама эволюция. Единицей эволюции
является популяция.
Виды,
населяющие Землю, образуют сообщества, пространственно-временные объединения,
одной из разновидностей которых является популяция.
К
основным характеристикам популяции относятся:
-
принадлежность особей к одному виду;
-
пространственно-временное единство;
-
сходство экологических потребностей;
- способность свободно и случайно скрещиваться между собой.
Поскольку
человек относится к одному из видов живых организмов, для него также характерно
образование популяций.
Замкнутые
популяции носят название изолятов.
Изоляты могут встречаться не только в географически изолированных местностях,
но и внутри крупных популяций, если какие-то группы людей по религиозным или
этническим причинам не смешиваются с остальной популяцией.
Среди
представителей любой популяции наблюдается определенная изменчивость, т. е.
разнообразие признаков у ее членов. Однако индивидуум не есть носитель всех расовых
свойств.
Расовая
изменчивость является групповой изменчивостью. Только в популяции
наследственная информация, пройдя апробацию на фенотипическом уровне
изменчивости, передается из поколения в поколение. Из мозаики популяционной
изменчивости складывается мозаика расовой изменчивости.
Представления
популяционной концепции расообразования опираются на достижения современной
эволюционной теории, популяционной генетики, новейшей биометрии.
Каждая популяция характеризуется определенным генофондом, т. е.
совокупностью аллелей, встречающихся у особей данной популяции.
Различия между популяциями определяются частотами генов и
генотипов. Если эволюционные процессы перестают ощутимо влиять на генофонд
популяции, то частота аллелей и генотипов определяется законом Харди–Вайнберга,
или законом генетической стабильности популяции.
Смысл этого закона заключается в том, что при определенных условиях
соотношение частот доминантных и рецессивных аллелей генов, сложившихся в
генофонде панмиксической популяции
(где свободно скрещиваются особи), сохраняется неизменным в ряду поколений.
В 1908 г. английский математик Дж. Харди и немецкий врач В.
Вайнберг независимо друг от друга установили, что частоты генотипов в популяции
можно выразить определенным математическим уравнением. Гены каждой пары, например
аллели А и а, распределяются таким образом, что отдельная особь, входящая в
состав популяции, может иметь один из трех генотипов – АА, Аа и аа. Для определения генетической
структуры панмиксической популяции необходимо помножить бином, члены которого
представляют частоты генов А и а для совокупности яйцеклеток (рА + qa), на другой такой же бином
для совокупности сперматозоидов (рА + qa). В
результате получим выражение, члены которого будут соответствовать частотам
различных генотипов в потомстве:
р2АА + 2рqАa + q2aа = (рА + qa)2,
где р и q –
частоты аллелей (А и а соответственно) аутосомного гена – р + q =1 (100 %); р2АА – частота встречаемости в популяции доминантных
гомозигот; 2 рqАa – частота гетерозигот; q2aа – частота рецессивных
гомозигот.
В случае трех аллелей (А1,
А2, А3) одного гена в популяции это будет разложение
квадрата трехчлена:
(рА1 + qА2 + rА3)2 = р2(А1
А1) + 2 рq(А1
А2) + 2 рr(А1 А3) +
+ q2(А2 А2)
+ 2 qr(А2 А3) + r2(А3
А3).
Чтобы рассчитать частоту генотипов для локусов с множественными
аллелями, используют формулу:
(р + q + r + …+ s)2 = 1,
где р, q, r …+ s – частоты
встречаемости различных аллелей.
Рассмотрим
замкнутую человеческую популяцию, в которой аллели А и а представлены тремя типами
сочетаний (АА, Аа и аа), и частота
встречаемости каждого из аллеля составляет
50 %, или 0,5. В такой популяции с равной вероятностью будут производиться
гаметы, несущие аллели А и аллель а, т.
е. частоты встречаемости этих гамет будут равны 0,5.
Для
изображения частот образования гомозигот и гетерозигот в следующем поколении
воспользуемся решеткой Пиннета (табл. 10).
Таблица 10
Решетка Пиннета
|
 ♀ ♀
♂
|
0,5 А
|
0,5 а
|
|
0,5А
0,5 а
|
0,25 АА
0,25 Аа
|
0,25 Аа
0,25 аа
|
Из
таблицы 10 видно, что в следующем поколении этой панмиксической популяции
соотношение частот аллелей А и а остается неизменным.
Установленная
Харди–Вайнбергом закономерность справедлива
для «идеальной» популяции, которая характеризуется:
- неограниченно большим числом особей, обеспечивающих
возможность свободного скрещивания;
- отсутствием мутационного процесса;
- отсутствием оттока какого-либо аллеля из генофонда
популяции за счет естественного отбора.
Популяций, отвечающих полностью требованиям закона Харди–Вайнберга,
в природе не существует. В каждой естественной популяции происходят
эволюционные процессы, связанные с изменением частот генов в результате:
- изменчивости;
- естественного отбора;
- потока и дрейфа генов.
Эти факторы играют важную роль в процессе расообразования.
Однако изменение частот аллелей под действием эволюционных факторов
осуществляется в популяциях очень медленно.
Популяционно-генетический метод позволяет понять направление
эволюции в истории человечества в разных частях мира.
К настоящему времени хорошо изучена частота распространения
аллелей системы группы крови АВО в
различных странах мира. Некоторые данные представлены в таблице 11 и на рисунке
118.
Таблица 11
Распространение
аллелей группы крови АВО
в
различных странах мира (%) (Айала и др., 1988)
|
Популяция
|
0
|
А
|
В
|
АВ
|
|
Русские
(Москва)
|
33,3
|
37,4
|
22,8
|
6,5
|
|
Французы
|
41,6
|
47,0
|
8,0
|
3,3
|
|
Китайцы
|
28,6
|
26,6
|
32,0
|
12,8
|
|
Японцы
|
30,1
|
38,4
|
21,9
|
9,7
|
|
Австралийцы
|
60,7
|
39,3
|
0
|
0
|
|
Бразилийцы
(Бороро)
|
100,0
|
0
|
0
|
0
|
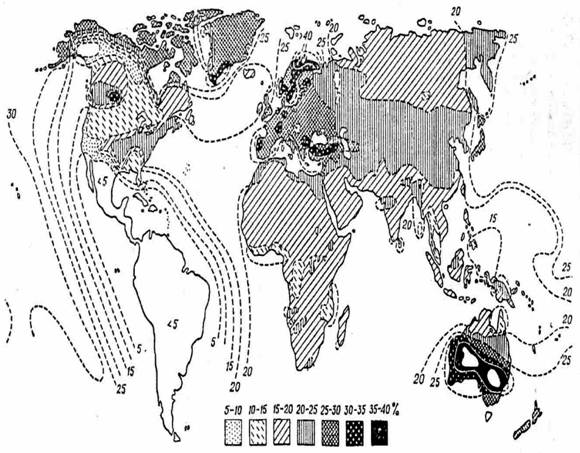
Рис. 118. Частота гена А групп
крови АВО в различных частях мира
(Mourant et. al., 1976)
Для
расчета частот аллелей в системе крови АВО
конечные формулы будут выглядеть следующим образом:
r = ; p =
; p =  – ; q =
– ; q = – ,
– ,
где А, В, О – частоты фенотипов; p – частота аллеля А;
q – частота аллеля В; r – частота аллеля О.
К основным факторам, порождающим изменчивость, относятся мутации, хромосомные перестройки и
рекомбинации.
Установление
с достаточной точностью факта появления признака в результате новой мутации у
человека представляет в каждом отдельном случае большие трудности. Для суждения
о частоте мутирования существуют косвенные методы, которые позволили по
отношению к человеку вычислить приблизительную частоту появления грубых мутаций
(табл. 12).
Таблица 12
Некоторые
наследственные метаболические дефекты
и
частоты встречаемости генотипов
|
Нарушение
|
Частота
рецессивного
гомозиготного
фенотипа (g2)
|
Частота
гетерозиготного
фенотипа (2рg)
|
|
Альбинизм
|
1 на 10000
|
1 на 50
|
|
Алкаптонурия
|
1 на 1000000
|
1 на 503
|
|
Семейная
амавротическая идиотия
|
1 на 40000
|
1 на 100
|
|
Сахарный диабет
|
1 на 200
|
1 на 8
|
|
Фенилкетонурия
|
1 на 10000
|
1 на 50
|
Для
проблемы расообразования весьма важно установление факта независимого появления
сходных (повторных) мутаций в разных группах населения. Такие признаки, как
альбинизм, короткопалость, карликовость, а из нормальных – рыжеволосость,
действительно возникли совершенно самостоятельно у отдельных индивидуумов
разных рас.
Еще
больший интерес представляют случаи появления независимых мутаций в
человеческой популяции, которые относятся к расовым признакам. Так появились
голубоглазость у субъектов африканских пигмеев, спирально закрученные волосы в
нидерландской семье.
Популяционно-генетический
метод позволяет представить какое-либо эволюционное событие в количественной
форме.
Ф.
Ливингстоном (1969 г.)
было выполнено моделирование эволюционного процесса с целью определения времени,
необходимого для установления различий по цвету кожи. Эксперимент проводился моделированием
процесса дифференциации с использованием ЭВМ. При этом сделаны достаточно
реалистичные предположения в свете того, что нам известно о наследовании цвета
кожи у человека и физиологических особенностях образования меланина:
1) цвет
кожи наследуется четырьмя несцепленными локусами по два аллеля в каждом;
2) из
четырех локусов два считались уменьшающими количество меланина (аналоги
ингибиторов), а два – увеличивающими его (аналоги
тирозиназы, катализирующей конверсию тирозина в меланин);
3)
аллели с главным эффектом (образование меланина и его угнетение считались
доминантными, так что для двух локусов доминировал «светлый» аллель, а для двух
– «темный»);
4)
рассматривались случаи без доминирования и с 80 % доминантностью.
Последние означают, что гетерозигота имеет фенотипическое значение, которое
является средней фенотипических значений обеих гомозигот и 80 % разницы между
этими значениями и доминантной гомозиготой (иными словами, фенотип гетерозиготы
очень близок доминантной гомозиготе);
5)
прямые и обратные мутации происходят с обычной частотой: 10–5 – 10–6;
6)
мутации включились в модель с одной и той же частотой от всех рецессивов к
доминантам и с другой, но тоже одинаковой, – в обратном направлении;
7)
количество меланина (степень пигментации определялась в балловой шкале от 0,0
до 8,0). Приспособленность каждой гаметной комбинации определялась как разница
между количеством меланина, продуцированного этой комбинацией, и фенотипом (оптимальный
фенотип – 8,0).
Главный
вывод этого эксперимента сводится к следующему: при 6 % различий в
приспособленности современная изменчивость цвета кожи могла быть достигнута за
800 поколений в отсутствии доминантности и через 1500 поколений при 80 %
доминантности. Даже при миграции 5 % между соседними изолятами в линейной их
последовательности возникают значительные различия в цвете кожи. Напомним, что
от верхнего палеолита нас отделяет около 2000 поколений, если принять
длительность поколения за 20 лет. Иными словами, современное типологическое
разнообразие в цвете кожи теоретически могло возникнуть после возникновения Homo sapiens sapiens.
4.3. Смешение, адаптация и изоляция как факторы
формирования рас
4.3.1. Смешение рас
Одним из
важных факторов формирования новых антропологических типов является смешение
кровей (метисация).
Для
смешения необходимо, чтобы две разные популяции встретились и хотя бы одна из
них была мигрантной. Понимание миграции в популяционной генетике связано с
представлением о «потоке» генов.
Метисация
популяции по средней арифметической величине, вычисленной для каждого
измерительного признака и для признака, определенного баллом, обычно занимает
промежуточное положение между смешивающимися расами.
Можно
выделить несколько областей наибольшего смешения рас в период до европейской
экспансии: Судан, Передняя Индия, Индо-Китай, Индонезия, Микронезия, Средняя
Азия, Западная Сибирь и Алтае-Саянское нагорье. После европейской экспансии наиболее
крупные масштабы смешение приняло в Южных штатах США, в Центральной и Южной
Америке.
На
настоящий момент в мире насчитываются многие сотни популяций смешанного
происхождения между значительно различающимися антропологическими типами. Антропологов
в первую очередь интересуют популяции, образовавшиеся при смешении европеоидов,
негроидов и монголоидов или некоторых других ясно различимых морфотипов (австралийские
аборигены, пигмеи и так далее).
Примером
промежуточности типа смешанной группы могут служить камчадалы, т. е. та группа русского населения Камчатки, которая
является в большей своей части, по крайней мере, по мужской линии, обрусевшими
потомками древних ительменов.
Антропологический
тип различных народов Западной Сибири, характеризующийся менее резко
выраженными чертами монголоидной расы, мог сложиться в процессе смешения
монголоидной и европеоидной рас.
Не
смешанное с коренными жителями русское старожильческое население Сибири
практически в полной мере сохранило антропологические особенности своих
исходных популяций (популяций – доноров).
Другой пример. Около 1 тыс. лет назад в
Юго-Западной Индии образовалась эндогамная группа потомков рабов арабских и
португальских купцов. Все эти рабы происходили из различных негроидных групп
Восточной Африки. Браков с индусами практически не было, так как этому
препятствовала кастовая система. Это небольшая группа (сейчас около 5 тыс.
чел.) по своим морфологическим чертам полностью негроидна, и по частотам генов различных групп крови, белков и ферментов
также ближе к своим африканским собратьям.
Потомки
второго поколения японских мигрантов в Бразилии не отличаются по описательным
признакам от японцев Японии, но имеют большие продольные размеры тела при
сокращении некоторых характерных пропорций (относительно длинный корпус при
укороченных ногах и так далее).
Для
решения вопроса о происхождении промежуточного антропологического типа важное
значение имеют данные по ископаемым типам на данной территории. Если по мере
углубления в древность промежуточный характер признаков исчезает и появляются
все более резко выраженные черты двух или более расовых комплексов, то всего
вероятнее, что ныне живущий промежуточный тип появился в результате их
смешения. Особенно отчетливо выступает смешанное происхождение какой-либо
группы в том случае, когда удается на археологическом материале установить
приход нового антропологического типа вместе с новой материальной культурой,
центр распространения которой известен в другой области. Наоборот, если
промежуточность просматривается до ранней поры позднего палеолита, то
значительно более вероятно считать именно промежуточный тип исходным, а крайние
варианты – результатом поздней дивергенции.
Примером
той роли, которую может сыграть ископаемый материал для разрешения этих
вопросов, является исследование черепов из Луговского могильника на территории
Татарстана. Он датируется периодом ананьинской культуры ранней поры железного
века. По антропологическим признакам черепа оказались монголоидного типа и
резко отличались от скифских и сарматских. Легкая монголоидность современных
народов, населяющих территорию Волго-Камья, несомненно, унаследована от ананьинцев,
смешавшихся позднее с различными европеоидными типами.
Примером
обратных результатов может послужить исследование мезолитских черепов Восточной
Африки. Оказалось, что в эпоху мезолита там не обнаружено резко выраженных
представителей ни негрской, ни европеоидной рас; черепа из пещеры Гэмбля
«нейтральны» по типу. Это подтверждение гипотезы, что происхождение
восточно-африканской расы нельзя сводить только к метисации, хотя последняя
широко распространена к югу от Эфиопии.
В
настоящее время существуют различные методы расчета смешения и генного потока.
Это так называемые расчеты доли примеси (или участия) исходных групп в смешанной.
Расчеты эти делаются по дискретным признакам с точно известной
наследственностью. Обычно это различные группы крови, белки и ферменты, а также
системы HL-A и Gm. Есть система подобного
расчета и для цвета кожи. В основе данных расчетов лежит предположение, что
частота гена в смешанной группе, пропорциональна вкладу исходных групп. Для
того чтобы избежать ошибок выборки, учитываются локусы (гены) с сильно
различающимися частотами аллелей в исходных группах, и результаты по разным
локусам усредняют. Основная формула следующая:
(1 – m)k =  ,
,
где g0 и Q – частоты генов в используемых
популяциях;
gk – частота генов в смешанной
популяции; m – доля генов в смешанной
популяции с частотой g0; k – число поколений.
Так как m рассматривается как поток генов в поколении, то
по дате начала смешения можно рассчитать среднюю скорость в поколении. Расчеты
этого показателя для смешанных групп США, Бразилии, Мексики, Камчатки дают
очень близкие цифры: от 2 до 5 %.
При
низкой интенсивности генного обмена (1–2 %) возрастает вероятность случайной
утери привнесенного нового аллеля в последовательном ряду поколений.
Отсюда
вывод, что должны существовать некие пороговые значения миграции, ниже которых
последствия генного потока не фиксируются. Исходя из экспериментальных расчетов
значения m, этот порог, скорее всего,
должен иметь значения ниже 1 %.
4.3.2. Адаптация и антропологические особенности
К
традиционным антропологическим темам относятся: изучение адаптационных
процессов, смешанных групп и наследования признаков в гибридных группах.
Ведущая теория расообразования рассматривает расовые типы как адаптационные.
Смешение
разных групп и адаптация у человека современного вида связаны с проблемой
миграций и мигрантных групп. В популяции, мигрировавшей в новые условия, возникают
процессы адаптации. Адаптогенез и отбор являются близкими понятиями, поскольку
адаптация рассматривается как причина отбора.
Эволюция
к большей адаптации с помощью отбора сразу по нескольким признакам – крайне медленный процесс, его
длительность измеряется десятками тысяч поколений, так как популяция не сможет
выдержать нужного для более быстрой эволюции числа так называемых «генетических
смертей» (этот эффект называется «дилеммой Холдейна»). Скорости становятся
более приемлемыми, если предположить, что интересующие антропологов признаки
наследуются вместе, т. е. сцепленно.
Для каждой среды обитания, в которой живет человек, характерен свой
климатический режим, который зависит от большого числа различных факторов:
географической широты, удаленности от моря, защищенности от ветров, рельефа
поверхности и высоты над уровнем моря.
Выше говорилось о значительном полиморфизме политипии человечества
по цвету кожи. С давних времен цвет кожи служил важным признаком расовой
дифференциации.
Существует взаимосвязь между территориальным распределением жителей
Земли по инсоляции и интенсивности пигментации кожи.
Важность цвета кожи в качестве расового признака объясняется связью
между солнечным освещением и продукцией витамина Д2, который
необходим для поддержания в организме баланса кальция. При недостатке витамина
Д в детском возрасте развивается рахит (нарушается процесс костеобразования
вследствие уменьшения Ca и P), снижается жизнеспособность и особенно
фертильность у женщин вследствие костных патологий тазового пояса.
Между тем количество витамина Д, синтезируемого в норме,
контролируется дозой солнечного облучения, которое проникает в клетки
дермального слоя. До периода, когда были разработаны способы искусственного
обогащения пищи витамином Д, люди были зависимы в продукции витамина Д2
от солнечного света. В высоких широтах при слабой инсоляции «выгодно» иметь
светлую кожу, т. к. определенное количество ультрафиолета необходимо для
образования витамина Д2. Посветление кожи у европейцев способствует
проникновению солнечного света глубоко в ткани человека, что ускоряет синтез
противорахитического витамина. Темная кожа имеет преимущества в широтах со
значительным количеством солнечных дней, т. к. она способна лучше воспринимать
тепловые лучи в инфракрасной части спектра от нагретых за день окружающих
предметов.
Миграция
людей с интенсивно пигментированной кожей к отдаленным от экватора широтам, а
людей с недостаточно пигментированной кожей – к тропическим широтам могла вести к дефициту
витамина Д у первых и избытку у вторых с соответствующими последствиями. Таким
образом, цвет кожи является селективным фактором эволюции.
С точки зрения
приспособления к климату большое значение имеют различия в характере
телосложения между разными популяциями. Эти различия подчиняются экологическим
правилам Бергмана и Аллена, которые справедливы для популяций животных и
человека. Правило Бергмана утверждает, что в пределах одного политипического
теплокровного вида размер тела подвида обычно увеличивается с уменьшением
температуры окружающей среды. Правило Аллена гласит: у теплокровных животных,
относящихся к одному виду, имеется тенденция к увеличению относительного размера
сильно выступающих частей (уши, хвост) с увеличением температуры окружающей
среды.
В ряде
исследований было показано, что форма и размеры тела человека также подчиняются
этим правилам.
В
популяциях жарких стран на всех континентах средний вес тела ниже, чем в
популяциях, живущих в умеренном и холодном климате. Было показано, что
отношение длины тела человека в сидячем положении к общей длине тела становится
меньше при увеличении средней годовой температуры, т. е. у жителей жарких стран
нижние конечности относительно длиннее. То же можно сказать и о верхних конечностях:
отношение размаха рук к длине тела у жителей жарких стран больше; у них меньше
размеры туловища. В совокупности все эти данные указывают на то, что при переходе
от умеренного к жаркому климату отношение веса тела к площади его поверхности
уменьшается. Корреляция между размерами или формой тела и средней температурой
тела объясняет до 50–60 % случаев межпопуляционной изменчивости.
Различие
в характере телосложения между коренастыми эскимосами, с одной стороны, и
длинными аборигенами Центральной Австралии или нилотами, с другой, служит примером
проявления правила Аллена. Правдоподобным кажется утверждение, что пигмеи и
нилоты представляют два крайних примера правил Бергмана и Аллена.
Несмотря
на скудность данных о толщине подкожного жирового слоя у представителей
различных народов, установлено, что у американских негров толщина кожной складки
меньше, чем у белых; у эскимосов жировой слой толще, чем у негров. У африканцев
эндоморфный компонент в конституции менее выражен, чем у европейцев и японцев.
В
зависимости от климатических условий проявляются некоторые особенности роста.
Тот факт, что удлиненная форма тела преобладает у людей, живущих в жарких
странах, согласуется с наблюдением, что период роста в этих странах удлинен, а
наступление половой зрелости несколько запаздывает. Удлиненная форма тела, т.
е. относительно большая длина на единицу веса, обычно связана с задержкой развития
скелета и общего физиологического созревания.
Существует
гипотеза, что черты лица монголоидного типа – это специальный приспособительный
признак для жизни в условиях суровых холодов. Для этого типа характерны
уменьшенные надбровные дуги и лобные пазухи, более широкие глазничные и
молярные области, уменьшенный носовой выступ. Сравнение происхождения монголоидной
расы и жителей континентальных областей Центральной Азии указывает, что особые
признаки монгольского глаза (узость щели, складка века, эпикантус) возникли как
защитный аппарат, охраняющий орган зрения от ветров, пыли и вредоносного
действия отраженной солнечной радиации на заснеженных пространствах.
Возникновение эпикантуса могло быть связано и с другими
причинами. Так, доказана внутригрупповая связь между выраженностью эпикантуса и
уплощенностью переносья – чем выше переносье, тем в среднем меньше эпикантус.
Эта связь обнаруживается у бурятов, киргизов, казахов, береговых чукчей,
эскимосов. Однако низкое переносье – не единственное и не достаточное условие
для возникновения эпикантуса.
Наличие
эпикантуса зависит также и от толщины жирового слоя под кожей верхнего века, и
до известной степени является «жировой» складкой верхнего века.
У части
туркмен Ашхабада с сильным жироотложением на лице эпикантус был отмечен
значительно чаще, чем у индивидов со слабой степенью жироотложения.
Усиленное
жироотложение на лице характерно для детей монголоидной расы, обладающей, как
известно, особенно сильным развитием эпикантуса.
Локальное
отложение жировой клетчатки у детей монголоидной расы могло иметь адаптивное
значение (предохраняло от обмерзания лица в условиях холодных зим) и, что менее
вероятно, является местным запасом питательного вещества с высоким калорийным
содержанием.
Стеатопигия
бушменов и готтентотов представляет также пример локального отложения жира у
населения, физический тип которого сформировался в условиях засушливого
климата.
Определенную
зависимость от климата испытывает и строение наружного носа. Не исключена
возможность, что крупные размеры и сильное выступание переднего носа имели известное
значение, там, где некоторая разреженность воздуха требовала большой площади
носового отверстия, а низкая температура благоприятствовала увеличению объема
носового входа как согревающей камеры. Подобные черты обнаруживаются у коренных
жителей Кавказа и переднеазиатских нагорий.
Ортогнантность европейцев и северных монголоидов могла
быть полезной, т. к. делала более крутым изгиб вдыхаемого воздуха и тем
предохраняла носоглотку от охлаждения.
4.3.3. Изоляция и расообразование
В числе
факторов расообразования, изоляция часто рассматривается как важный, а иногда
единственный фактор политипии. Данные
зоогеографии свидетельствуют о том, что величина различий между подвидами
зависит от степени и длительности их изолированного существования и отсутствия
смешений между ними. Первобытное общество прошло через очень продолжительную
стадию существования изолированных коллективов или совокупности коллективов, связанных
и ограниченных рамками эндогамии. Причины изоляции могут быть разными, но с
биологической точки зрения становление изолятов связано с отсутствием генного
потока и миграцией извне.
Интенсивность
воздействия изоляции на межгрупповую изменчивость прямо пропорциональна времени
и обратно пропорциональна численности популяции. Первое обусловлено постоянством
частоты мутаций, их случайностью и малой вероятностью закрепления в популяции
мутантной формы аллеля, если его не поддерживает отбор. В силу этого изоляты
большой численности дивергируют медленно. В малых изолятах генетический дрейф
может способствовать быстрому изменению генофонда за счет случайного закрепления
одних аллелей и потери других.
Одним из
последствий малочисленности популяций является инбридинг, т. е. увеличение
вероятности браков между близкими родственниками. Обычно он приводит к росту гомозиготности.
Так как многие наследственные дефекты рецессивны, то увеличивается вероятность
фенотипических отклонений от нормы. Все это может приводить к увеличению числа
людей с контрастными фенотипами и к уменьшению генетического разнообразия (за
счет потери аллелей), снижению гетерозиготности и общего фенотипического
разнообразия. В известной степени изоляцию можно уподобить центрифугированию, в
котором скорость фракционирования пропорциональна инбридингу.
Следует
подчеркнуть, что в отсутствие «вредных» аллелей сам факт увеличения
гомозиготности, видимо, не наносит заметного ущерба популяции. Существует также
мнение, что при некоторых обстоятельствах (например, при пониженной
жизнеспособности гетерозигот и отсутствии воспроизводства у одной из гомозигот)
инбридинг будет снижать частоту неблагоприятного аллеля.
Ранее мы
отмечали, что Homo sapiens в течение длительного
периода состоял преимущественно из небольших по численности популяций.
Несомненно, некоторые из них могли выпадать из системы генного обмена, т. е.
становились изолятами. Время изоляции могло быть большим. Например, для
населения Австралии и обеих Америк практическое отсутствие генного обмена с
Евразией длилось тысячелетиями. В таком же положении оказывались и некоторые
окраинные группы Старого Света. Климатогеографические предпосылки для этого
были.
Косвенным
подтверждением роли изоляции и сопряженных с нею случайных процессов в создании
политипии является значительное сходство филогенетических «древов» традиционных
классификаций с современными нумерическими классификациями, в большинстве
которых делается предположение о ведущей роли генетического дрейфа.
Есть еще одна особенность изоляции как фактора эволюции.
В изолятах могут длительное время сохраняться реликтовые формы. Правда,
длительное существование малочисленного изолята – событие маловероятное.
Случайная гибель нескольких особей может фатально нарушить демографическую
структуру, и популяция вымрет. Можно полагать, что это неоднократно происходило
в истории человечества. К такому же результату приводит уменьшение генетической
изменчивости, приводящее к сужению нормы реакции, а соответственно и
адаптационных способностей индивидов и популяций.
Контрольные вопросы
1. Различие
между понятиями нация и раса.
2. Многообразие определений понятия раса.
3. История научных классификаций рас.
4. Современная классификация рас.
5. Характеристика европеоидной большой расы.
6. Антропологические особенности малых рас,
входящих в европеоидную расу.
7. Антропологические особенности монголоидной
расы.
8. Характеристика малых рас в составе
монголоидной расы.
9. Большая австрало-негроидная, или
экваториальная, раса.
10. Деление экваториальной расы на малые расы и
их особенности.
11. Причины и факторы образования рас.
12. Понятие о качественных и количественных
признаках.
13. Основные характеристики популяций.
14. Закон генетической стабильности популяции.
15. Причины изменения частот генов популяции.
16. Использование популяционно-генетического
метода в изучении направлений эволюций.
17. Факты независимого появления сходных
мутаций у индивидов сходных рас.
18. Подходы
к моделированию эволюционного процесса.
19. Смешение как фактор формирования новых антропологических
типов.
20. Археологический материал в решении вопроса
о происхождении промежуточных антропологических типов.
21. Современные методы расчета смешения и
генного потока.
22. Роль
адаптационных процессов в расообразовании.
23.
Правило Бергмана и Аллена.
24. Возможные причины возникновения эпикантуса,
стеатопигии, наружного уха и других антропологических признаков.
25. Роль
изоляции в создании политипии.