Тема 8. НЕРВНАЯ ТКАНЬ
План:
2. Глия.
4. Синапсы. Нервные окончания.
5. Понятие о рефлекторной дуге.
Нервная ткань - это система взаимосвязанных нервных клеток и нейроглии, обеспечивающих специфические функции восприятия раздражений, возбуждения, выработки импульса и передачи его. Она является основой строения органов нервной системы, обеспечивающих регуляцию всех тканей и органов, их интеграцию в организме и связь с окружающей средой.
Нервные клетки
(нейроны - neuronum, нейроциты)
- основные структурные компоненты нервной ткани, выполняющие специфическую
функцию. Нейроглия (neuroglia) обеспечивает
существование и функционирование нервных клеток, осуществляя опорную,
трофическую, разграничительную, секреторную и защитную функции.
1.
Нейроны
Нейроны,
или нейроциты - специализированные клетки
нервной системы, ответственные за рецепцию, обработку (процессинг) стимулов,
проведение импульса и влияние на другие нейроны, мышечные или секреторные клетки.
Нейроны выделяют нейромедиаторы и другие вещества,
передающие информацию. Нейрон является морфологически и функционально
самостоятельной единицей, но с помощью своих отростков осуществляет синаптический контакт с другими
нейронами, образуя рефлекторные дуги - звенья цепи, из которой
построена нервная система. В зависимости от функции в рефлекторной дуге
различают рецепторные (чувствительные, афферентные), ассоциативные и эфферентные (эффекторные) нейроны. Афферентные нейроны воспринимают
импульс, эфферентные передают его на ткани рабочих органов, побуждая их к
действию, а ассоциативные осуществляют связь между нейронами.
Нейроны отличаются большим разнообразием форм и
размеров. Диаметр тел клеток-зерен коры мозжечка 4-6 мкм, а гигантских пирамидных
нейронов двигательной зоны коры большого мозга - 130-150 мкм. Обычно нейроны
состоят из тела (перикариона)
и отростков: аксона и различного числа ветвящихся
дендритов (рис. 48).

Рис. 48. Нейрон.
1 - дендриты, 2 - перикарион,
3- аксонный холмик, 4 – аксон, 5- шванновская
клетка, 6- ядро шванновской клетки, 7 – перехват Ранвье, 8 – терминали.
По количеству отростков различают
униполярные нейроны, имеющие только аксон (у высших животных и человека
обычно не встречаются), биполярные, имеющие аксон и один
дендрит, и мультиполярные,
имеющие аксон и много дендритов (рис. 49). Иногда среди биполярных нейронов
встречается псевдоуниполярный,
от тела которого отходит один общий вырост - отросток,
разделяющийся затем на дендрит и аксон. Псевдоуниполярные
нейроны присутствуют в спинальных ганглиях, биполярные - в органах чувств.
Большинство нейронов мультиполярные. Их формы
чрезвычайно разнообразны. Аксон и
его коллатерали оканчиваются, разветвляясь на несколько веточек, называемых телодендронами (telodendron),
последние заканчиваются терминальными утолщениями.
В нейроне различают часть, специализированную на
рецепции стимулов, дендриты и тело - перикарион,
трофическую часть (тело нейрона) и проводящую, передающую импульс (аксон).
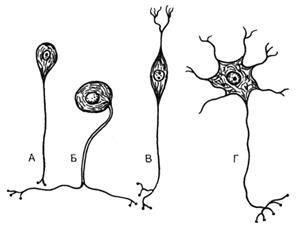
Рис. 49. Типы нервных клеток
(схема по Т.Н.Радостиной, Л.С.Румянцевой).
А - униполярный нейрон; Б - псевдоуниполярный
нейрон; В - биполярный нейрон; Г - мультиполярный
нейрон.
соответствии с высокой активностью метаболизма нейроцитов
хроматин в их ядрах диспергирован. В ядре имеется 1, а иногда 2-3 крупных ядрышка. Усиление функциональной активности нейронов
обычно сопровождается увеличением объема (и количества) ядрышек.
Дендриты
представляют собой истинные выпячивания тела клетки. Они содержат те же
органеллы, что и тело клетки: глыбки хроматофильной субстанции (гранулярный эндоплазматический ретикулум и полисомы),
митохондрии, большое количество нейротубул
(микротрубочек) и нейрофила-ментов. За счет дендритов
рецепторная поверхность нейрона увеличивается в 1000 и более раз. Так, дендриты
грушевидных нейронов (клеток Пурки-нье) коры мозжечка
увеличивают площадь рецепторной поверхности от 250 мкм2
до 27 000 мкм2, и на поверхности этих клеток обнаруживается до 200
000 синаптических окончаний.
Аксон - отросток,
по которому импульс передается от тела клетки. Он содержит митохондрии, нейротубулы и нейрофиламенты, а
также агранулярный эндо плазматический (но не
гранулярный) ретикулум.
Ядро нейрона. Подавляющее большинство нейронов человека содержит
одно ядро, расположенное чаще в центре, реже - эксцентрично. Двуядерные и тем
более многоядерные нейроны встречаются крайне редко. Исключение составляют
нейроны некоторых ганглиев вегетативной нервной системы; например, в
предстательной железе и шейке матки иногда встречаются нейроны, содержащие до
15 ядер.
Плазмолемма нейрона обладает способностью генерировать и проводить
импульс. Ее интегральными белками являются белки, функционирующие как
ионно-избирательные каналы, и рецепторные белки, вызывающие реакции нейронов на
специфические стимулы. Ионные каналы могут быть открыты, закрыты или
инактивированы. В покоящемся нейроне мембранный потенциал покоя равен -60-70
мВ. Потенциал покоя создается за счет выведения Na+
из клетки. Большинство Na+- и К+-каналов
при этом закрыты. Переход каналов из закрытого состояния в открытое
регулируется мембранным потенциалом.
Хроматофильная субстанция (тигроид, или тельца Ниссля) – это хорошо развитый гранулярный
эндоплазматический ретикулум; она синтезирует
нейросекреторные белки, интегральные белки плазмолеммы и белки лизосом. Для
аксонов, не имеющих органелл, синтезирующих белок, характерен постоянный ток
цитоплазмы от перикариона к терминалям
со скоростью 1-3 мм в сутки.
Аппарат Гольджи в нейронах хорошо развит. Пузырьки аппарата Гольджи транспортируют белки, синтезированные в гранулярном эндоплазматическом ретикулуме
либо к плазмолемме (интегральные белки), либо в терминали
(нейропептиды, нейросекрет), либо в лизосомы (лизосомальные гидролазы и мембраны лизосом).
Митохондрии обеспечивают энергией такие процессы, как транспорт
ионов и синтез белков. Нейроны нуждаются в постоянном притоке глюкозы и
кислорода с кровью, и прекращение кровоснабжения головного мозга вызывает
потерю сознания.
Лизосомы участвуют в ферментативном расщеплении компонентов
клетки рецепторов и мембран, часть из которых может рециркулировать.
Возрастные изменения нейронов сопровождаются
накоплением липофусцина, разрушением крист
митохондрий. Липофусцин - "пигмент старения" - желто-бурого
цвета липопротеидной природы, представляющий собой остаточные тельца (телолизосомы) с продуктами непереваренных структур.
Из элементов цитоскелета
в цитоплазме нейронов присутствуют нейрофиламенты
диаметром 12 нм и нейротубулы
диаметром 24-27 нм, которые участвуют в поддержании
формы клеток, росте отростков и аксональном транспорте.
Способность синтезировать и секретировать биологически
активные вещества, в частности медиаторы (ацетилхолин, норадреналин, серотонин
и др.), свойственна всем нейроцитам. Однако
существуют нейроциты, специализированные
преимущественно для выполнения этой функции, - секреторные
нейроны, например клетки нейросекреторных ядер гипоталамической области
головного мозга. Секреторные нейроны имеют ряд специфических морфологических
признаков. Это крупные нейроны. Хроматофильная
субстанция преимущественно располагается по периферии тела клеток. В цитоплазме
нейронов и в аксонах находятся различной величины гранулы секрета нейросекрета
(substantia neurosecretoria),
содержащие белок, а в некоторых случаях липиды и полисахариды. Гранулы
нейросекрета выводятся в кровь или мозговую жидкость. Многие секреторные
нейроны имеют ядра неправильной формы, что свидетельствует об их высокой
функциональной активности. Нейросекреты выполняют роль
нейрорегуляторов, участвуя во взаимодействии нервной
и гуморальной систем интеграции.
2.
Нейроглия
Нейроны - высокоспециализированные клетки,
существующие и функционирующие в строго определенной среде. Такую среду им обеспечивает
нейроглия. Нейроглия выполняет следующие функции: опорную, трофическую,
разграничительную, поддержание постоянства среды вокруг нейронов, защитную,
секреторную.
Глия центральной
нервной системы. Клетки глии центральной
нервной системы делятся на макроглию (глиоциты) и микроглию.
Макроглия развивается из глиобластов
нервной трубки. К макроглии относятся эпендимоциты, астроциты и олигодендроглиоциты (рис. 50, 51).
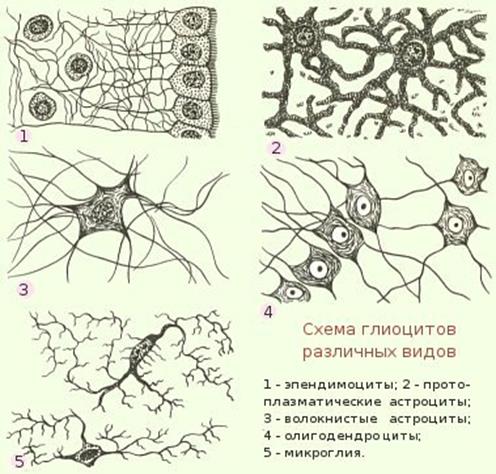
Рис. 50. Клетки нейроглии.
Эпендимоциты выстилают
желудочки головного мозга и центральный канал спинного мозга. Эти клетки
цилиндрической формы. Они образуют слой типа эпителия. Между соседними клетками
имеются щелевидные соединения и пояски сцепления, но плотные соединения
отсутствуют, так что цереброспинальная жидкость может проникать между ними в
нервную ткань. Большинство эпендимоцитов имеют
подвижные реснички, вызывающие ток цереброспинальной жидкости. Базальная
поверхность большинства эпендимоцитов ровная, но
некоторые клетки имеют длинный отросток, идущий глубоко в нервную ткань, и
почти лишены ресничек. Такие клетки называются таницитоми.
Они многочисленны в дне III желудочка. Считается, что эти клетки передают
информацию о составе цереброспинальной жидкости на первичную капиллярную сеть
воротной системы гипофиза. Эпендимный эпителий
сосудистых сплетений желудочков продуцирует цереброспинальную жидкость.
Цитоплазма эпендимоцитов содержит многочисленные
митохондрии, аппарат Гольджи, расположенный над ядром
и слаборазвитый гранулярный эндоплазматический ретикулум.
Астроциты (astrocyti, от греч. astron - звезда, kytos - клетка) - клетки отростчатой
формы, бедные органеллами. Они выполняют в основном опорную и разграничительную
функции (см. рис. 50, 51). Различают протоплазматические астроциты,
локализующиеся в сером веществе центральной нервной системы, и волокнистые астроциты, присутствующие в белом веществе. Протоплазматические астроциты
характеризуются короткими сильно ветвящимися отростками и светлым сферическим
ядром. Волокнистые астроциты имеют 20-40 длинных, слабо ветвящихся отростков,
в которых много фибрилл, состоящих из промежуточных филаментов
диаметром 10 нм. В филаментах
выявляется глиальный фибриллярный кислый белок. Отростки астроцитов
тянутся к базальным мембранам капилляров, к телам и дендритам нейронов, окружая
синапсы и, отделяя их друг от друга, а также к мягкой
мозговой оболочке, образуя пиоглиальную мембрану,
граничащую с субарахноидальным пространством. Подходя к капиллярам, их отростки
образуют расширенные "ножки", полностью окружающие сосуд. Астроциты накапливают и передают вещества от капилляров к
нейронам, захватывают избыток экстрацеллюлярного
калия и других веществ, таких как нейромедиаторы, из экстрацеллюлярного пространства после интенсивной нейрональной активности.
Олигодендроциты имеют более мелкие по сравнению с астроцитами
и более интенсивно окрашивающиеся ядра. Их отростки немногочисленны. Олигодендроглиоциты присутствуют как в сером, так и в белом
веществе. В сером веществе они локализуются вблизи перикарионов.
В белом веществе их отростки образуют миелиновый слой в миелиновых нервных
волокнах, причем в противоположность нейролеммоцитам
периферической нервной системы один олигодендроглиоцит
может участвовать в миелинизации нескольких аксонов
(рис. 51). Один отросток формирует миелиновый слой одного интернодального
сегмента. Цитоплазма олигодендроцитов электронноплотная, содержит много митохондрий, развитый
аппарат Гольджи, цистерны гранулярного
эндоплазматического ретикулума, многочисленные
микротрубочки.
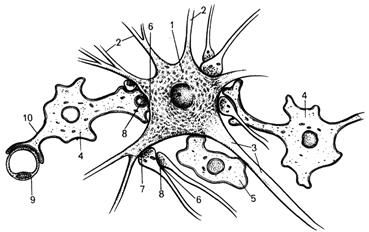
Рис. 51. Взаимоотношения
нейрона, астроглии, олигодендроглии
и нервных терминалей (по Г.Р.Нобаку, Н.Л.Стромингеру, Р.Д.Демаресту).
1 - тело нейрона; 2 - дендриты; 3 - аксон; 4 - астроглиоциты;
5 - олигодендроцит; 6 - аксоаксональный
синапс; 7 - аксодендритический синапс; 8 - аксосоматический синапс; 9 -
капилляр; 10 - периваскулярная ножка астроцита.
Микроглия представляет собой фагоцитирующие
клетки из системы мононуклеарных фагоцитов и
происходящие из стволовой кроветворной клетки. Ее функция - защита от инфекции
и повреждения и удаление продуктов разрушения нервной ткани. Клетки микроглии характеризуются небольшими размерами, телами
продолговатой формы, обладают слабой фагоцитарной активностью. Ветвистая микроглия
встречается как в сером, так и в белом веществе центральной нервной системы. В
развивающемся мозгу млекопитающих обнаруживается временная форма микроглии - амебоидная микроглия.
Клетки амебоидной микроглии формируют филоподии и складки плазмолеммы. В их цитоплазме
присутствуют многочисленные фаголизосомы и
пластинчатые тельца. Амебоидные тельца отличаются высокой активностью лизосомальных ферментов. Активно фагоцитирующая
амебоидная микроглия необходима
в раннем постнатальном периоде, когда гематоэнцефалический барьер еще не вполне
развит и вещества из крови легко попадают в центральную нервную систему.
Считают также, что она способствует удалению обломков клеток, появляющихся в
результате запрограммированной гибели избыточных нейронов и их отростков в
процессе дифференцировки нервной системы. Полагают, что, созревая, амебоидные микроглиальные клетки превращаются в ветвистую
микроглию.
Реактивная микроглия появляется после травмы в любой области мозга. Она не
имеет ветвящихся отростков, как покоящаяся микроглия,
не имеет псевдоподий и филоподии, как амебоидная микроглия. В цитоплазме клеток реактивной микроглии присутствуют плотные тельца, липидные включения,
лизосомы. Есть данные о том, что реактивная микроглия
формируется вследствие активации покоящейся при травмах центральной нервной
системы.
Глия
периферической нервной системы
(периферическая нейроглия) в отличие от макроглии
центральной нервной системы происходит из нервного гребня. К периферической
нейроглии относятся нейролеммоциты
(шванновские клетки) и глиоциты ганглиев (мантийные глиоциты).
Нейролеммоциты формируют оболочки отростков нервных клеток в нервных
волокнах периферической нервной системы. Глиоциты ганглиев окружают тела нейронов в нервных узлах и
участвуют в обмене веществ нейронов.
3.
Нервные волокна
Отростки нервных клеток, покрытые оболочками,
называются нервными волокнами. По
строению оболочек различают миелиновые и безмиелиновые
нервные волокна (рис. 52, ). Отросток нейрона в
нервном волокне называют осевым цилиндром, или аксоном, так
как чаще всего (за исключением чувствительных нервов) в составе нервных волокон
находятся именно аксоны.
В центральной нервной системе оболочки отростков
нейронов образуют отростки олигодендроглиоцитов, а в
периферической - нейролеммоциты.
Безмиелиновые нервные волокна находятся преимущественно в составе вегетативной нервной системы. Нейролеммоциты оболочек безмиелиновых
нервных волокон, располагаясь плотно, образуют тяжи, в которых на определенном
расстоянии друг от друга видны овальные ядра.
|
|
|
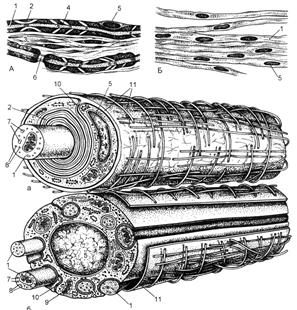
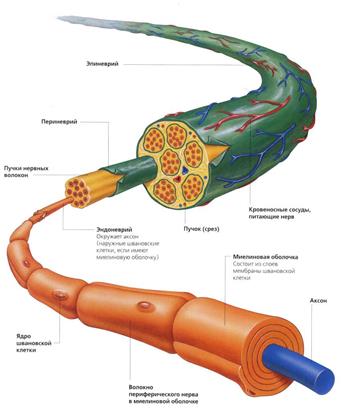
Рис. 52.
Строение нервных волокон на светооптическом (А, Б) и ультрамикроскопическом (а,
б) уровнях (схема по Т.Н.Радостиной, Ю.И.Афанасьеву, Л.С.Румянцевой).
А, а - миелиновое волокно; Б, б - безмиелиновое
волокно; 1 - осевые цилиндры; 2 - миелиновый слой; 3 - соединительная ткань; 4
- насечка миелина; 5 - ядро нейролеммоцита; 6 -
узловой перехват; 7 - микротрубочки; 8 - нейрофиламенты;
9 - митохондрии; 10 - мезаксон; 11 - базальная
мембрана.
В нервных волокнах внутренних органов, как правило, в таком
тяже имеется не один, а несколько (10-20) осевых цилиндров, принадлежащих
различным нейронам. Они могут, покидая одно волокно, переходить в смежное. Такие волокна, содержащие несколько осевых
цилиндров, называются волокнами кабельного типа. При электронной
микроскопии безмиелиновых нервных волокон видно, что
по мере погружения осевых цилиндров в тяж неиролеммоцитов
оболочки последних прогибаются, плотно охватывают осевые
цилиндры и, смыкаясь над ними, образуют глубокие складки, на дне которых и
располагаются отдельные осевые цилиндры. Сближенные в области складки участки
оболочки нейролеммоцита образуют сдвоенную мембрану -
мезаксон, на которой как бы подвешен осевой
цилиндр (см. рис. 52). Оболочки нейролеммоцитов очень
тонкие, поэтому ни мезаксона, ни границ этих клеток
под световым микроскопом нельзя рассмотреть, и оболочка безмиелиновых
волокон в этих условиях выявляется как однородный тяж цитоплазмы,
"одевающий" осевые цилиндры.
Миелиновые нервные волокна встречаются как в центральной, так и в периферической
нервной системе. Они значительно толще безмиелиновых
нервных волокон. Диаметр поперечного сечения их колеблется от 2 до 20 мкм. Они
также состоят из осевого цилиндра, "одетого"
оболочкой из нейролеммоцитов (шванновских клеток), но диаметр осевых цилиндров этого типа
волокон значительно толще, а оболочка сложнее. В сформированном миелиновом
волокне принято различать два слоя оболочки: внутренний, более толстый, - миелиновыи слой, а) и наружный, тонкий, состоящий из цитоплазмы, ядер нейролеммоцитов и нейролеммы.
В процессе развития аксон погружается в желобок на
поверхности нейролеммоцита. Края желобка смыкаются.
При этом образуется двойная складка плазмолеммы нейролеммоцита
- мезаксон. Мезаксон удлиняется, концентрически наслаивается на осевой
цилиндр и образует вокруг него плотную слоистую зону -
миелиновый слой (рис. 53).
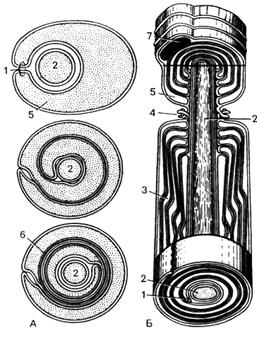
|
Рис. 53. Развитие и строение
миелинового волокна (схема). 1 - дубликация оболочки
нейролеммоцита (мезаксон);
2 - аксон; 3 -насечка миелина; 4 - пальцевидные контакты нейролеммоцита
в области перехвата; 5 - цитоплазма нейролеммоцита;
6 - спирально закрученный мезаксон (миелин); 7 -
ядро нейролеммоцита. |
На электронных микрофотографиях видны главные плотные
и интрапериодальные линии. Отсутствие миелинового
слоя в области узловых перехватов объясняется тем, что в этом участке волокна
кончается один нейролеммоцит и начинается другой.
Осевой цилиндр в этом месте частично прикрыт интердигитирующими
отростками нейролеммоцитов. Аксолемма
(оболочка аксона) обладает в области перехвата значительной электронной
плотностью. Наличие большого числа митохондрий в этой области свидетельствует о
высокой метаболической активности аксолеммы. Следует
отметить, что ветвление аксонов происходит также в области перехватов.
Рис. 54. Строение нерва.
Миелиновые волокна центральной нервной системы отличаются тем, что в них миелиновый слой формирует
один из отростков олигодендроглиоцита. Остальные его
отростки участвуют в образовании миелинового слоя других миелиновых волокон
(каждый в пределах одного межузлового сегмента).
Миелиновые волокна центральной нервной системы не имеют насечек миелина, а
нервные волокна не окружены базальными мембранами.
Скорость передачи импульса миелиновыми волокнами
больше, чем безмиелиновыми. Тонкие волокна, бедные
миелином, и безмиелиновые волокна проводят нервный
импульс со скоростью 1-2 м/с, тогда как толстые миелиновые - со скоростью 5-120
м/с.
Разрешение миелиновой оболочки нарушает проводимость
нервных импульсов и работу нервной системы и организма, вплоть до паралича.
В безмиелиновом
волокне волна деполяризации мембраны идет по всей аксолемме,
не прерываясь, а в миелиновом возникает только в области перехвата. Таким образом, для миелиновых волокон характерно сальтаторное проведение возбуждения, т.е. прыжками. Между
перехватами идет электрический ток, скорость которого выше, чем прохождение
волны деполяризации по аксолемме.
4.
Синапсы. Нервные окончания
Нервные волокна заканчиваются концевыми аппаратами - нервными
окончаниями. Различают 3 группы нервных окончаний: концевые аппараты,
образующие межнейрональные синапсы и осуществляющие
связь нейронов между собой; эффекторные окончания
(эффекторы), передающие нервный импульс на ткани рабочего органа; рецепторные (аффекторные, или чувствительные).
Синапсы -
это межклеточные контакты нервной ткани, предназначенные для передачи импульса
с одного нейрона на другой или на мышечные и железистые структуры. Синапсы
обеспечивают поляризацию проведения импульса по цепи нейронов, т.е.
определяют направление проведения импульса. Если раздражать аксон электрическим
током, импульс пойдет в обоих направлениях, но импульс, идущий в сторону тела
нейрона и его дендритов, не может быть передан на другие нейроны. Только
импульс, достигающий терминален аксона, с помощью
синапсов может передать возбуждение на другой нейрон, мышечную или железистую
клетку.
В зависимости от способа передачи импульса синапсы
могут быть химическими или электрическими (электротоническими).
В зависимости от локализации окончаний терминальных
веточек аксона первого нейрона различают аксодендритические, аксосоматические и
аксоаксональные синапсы.
Химические синапсы передают импульс на другую клетку с помощью специальных биологически
активных веществ - нейромедиаторов, находящихся в синаптических пузырьках (рис. 55). Терминаль
аксона представляет собой пресинаптическую
часть, а область второго нейрона, или другой иннервируемой клетки, с
которой она контактирует, - постсинаптическую часть.
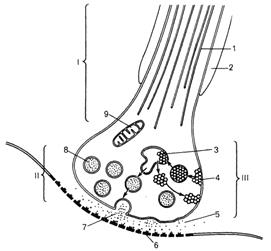
Рис. 55. Циклические
изменения синаптических пузырьков в синапсе (схема по
Ч.Р.Нобаку, Н.Л.Стромингеру,
Р.Дж.Демаресту).
I - нервное волокно; II - синапс; III - пресинаптическая часть; 1 - микротрубочки; 2 -миелиновая
оболочка; 3 - формирование цистерн, из которых вновь образуются синаптические пузырьки; 4 - образование новых мембран синаптических пузырьков путем пиноцитоза
(эндоцитоза) порций нейротрансмиттера;
5 - синаптическая щель; 6 - постсинаптическая
мембрана; 7 - слияние мембраны синаптического
пузырька с плазмолеммой и высвобождение нейротрансмиттера
путем экзоцитоза в синаптическую
щель; 8 - синаптические
пузырьки; 9 - митохондрии.
В пресинаптической части
находятся синаптические пузырьки, многочисленные
митохондрии и отдельные нейрофиламенты. Форма и
содержимое синаптических пузырьков связаны
с функцией синапса. Например, округлые прозрачные пузырьки диаметром 30-50 мн присутствуют в синапсах, где передача импульса
совершается с помощью ацетилхолина (холинергические синапсы).
Холинергическими являются парасимпатические и преганглионарные
симпатические синапсы, аксо-мышечные синапсы (см. ниже) и некоторые синапсы центральной
нервной системы. В синапсах, в которых в качестве нейромедиатора
используется норадреналин (адренергические синапсы), имеются синаптические пузырьки диаметром 50-90 нм
с плотной сердцевиной диаметром 15-25 нм.
Норадреналин является медиатором постганглионарных
симпатических синапсов. Адетилхолин и норадреналин -
наиболее распространенные медиаторы, но существует и множество
других. Различают низкомолекулярные, т.е. с небольшой относительной
молекулярной массой, нейромедиаторы (ацетилхолин,
норадреналин, дофамин, серотонин, глицин, гамма-аминомасляная кислота,
серотонин, гистамин, глютамат) и нейропептиды:
опиоидные (эндорфины, энкефалины,
динорины), вещество Р и др.
Дофамин, глицин и гамма-аминомасляная
кислота являются медиаторами тормозящих синапсов. Вырабатывающиеся в головном
мозге эндорфины и энкефалины
являются ингибиторами восприятия боли. Однако большинство медиаторов и
соответственно синапсов являются возбуждающими.
Область синаптического
контакта между двумя нейронами состоит из пресинаптической
мембраны, синаптической щели и постсинаптической
мембраны. Пре- и постсинаптическая мембраны кажутся
утолщенными из-за скопления под плазмолеммами электронноплотных
белков цитоплазмы, формирующих синапс. Цитоплазматические
филаменты, подобно филаментам
десмосом, прикрепляются к мембранам изнутри.
Пресинаптическая мембрана - это
мембрана клетки, передающей импульс (аксолемма). В
этой области локализованы кальциевые каналы, способствующие слиянию синаптических пузырьков с пресинаптической
мембраной и выделению медиатора в синаптическую щель.
Синоптическая щель между пре- и
постсинаптической мембранами имеет ширину 20-30 нм.
Мембраны прочно прикреплены друг к другу в си-наптической
области филаментами, пересекающими
синаптическую щель.
Постсинаптическая мембрана - это участок плазмолеммы клетки, воспринимающий
медиаторы генерирующий импульс. Она снабжена рецептор-ными зонами для восприятия соответствующего нейромедиатора.
В целом процессы в синапсе происходят в следующем
порядке:
1) волна деполяризации доходит до пресинаптической
мембраны;
2) открываются кальциевые каналы, и Са2+ входит в терминаль;
3) вхождение Са2+
в терминаль вызывает экзоцитоз
нейромедиатора; при этом мембрана синаптических
пузырьков входит в состав пресинаптической мембраны,
а медиатор попадает в синаптическую щель; в
дальнейшем мембраны синаптических пузырьков, вошедшие
в со став пресинаптической мембраны, и часть
медиатора подвергаются эндоцитозу
и происходит рециркуляция синаптических пузырьков
(рис. 55), а часть мембран и нейромедиатора с помощью
ретроградного транспорта поступает в перикарион и
разрушается лизосомами;
4) нейромедиатор
диффундирует через синаптическую щель и связывается с
рецепторными участками на постсинаптической мембране, что вызывает молекулярные
изменения в постсинаптической мембране, приводящие к 5) открытию ионных каналов
и 6) созданию постсинаптических потенциалов, обусловливающих реакции
возбуждения или торможения.
Электрические, или электротонические,
синапсы в нервной системе
млекопитающих встречаются относительно редко. В области таких синапсов
цитоплазмы соседних нейронов связаны щелевидными соединениями (контактами),
обеспечивающими прохождение ионов из одной клетки в другую, а
следовательно, электрическое взаимодействие этих клеток. Эти синапсы
способствуют синхронизации активности.
Эффекторные нервные окончания
Эффекторные нервные окончания бывают двух типов - двигательные и
секреторные.
Двигательные нервные окончания - это концевые аппараты аксонов двигательных клеток
соматической, или вегетативной, нервной системы. При их участии нервный импульс
передается на ткани рабочих органов. Двигательные окончания в
поперечнополосатых мышцах называются нервно-мышечными окончаниями.
Они представляют собой окончания аксонов клеток двигательных
ядер передних рогов спинного мозга или моторных ядер головного мозга.
Нервно-мышечное окончание состоит из концевого ветвления осевого цилиндра
нервного волокна и специализированного участка мышечного волокна (рис. 56).
Миелиновое нервное волокно, подойдя к мышечному волокну, теряет миелиновый слой
и погружается в него, вовлекая за собой его плазмолемму и базальную мембрану.
Мембрана мышечного волокна
образует многочисленные складки, формирующие вторичные синаптические
щели эффекторного окончания. В области окончания
мышечное волокно не имеет типичной поперечной исчерченности
и характеризуется обилием митохондрий, скоплением круглых или слегка овальных
ядер. Саркоплазма с митохондриями и ядрами в совокупности образует постсинаптическую
часть синапса.
Терминальные ветви нервного волокна в мионевральном
синапсе характеризуются обилием митохондрий и многочисленными пресинаптическими пузырьками, содержащими
характерный для этого вида окончаний медиатор - ацетилхолин. При возбуждении
ацетилхолин поступает через пресинаптическую
мембрану в синаптическую щель на холинорецепторы постси-наптической
(мышечной) мембраны, вызывая ее возбуждение (волну деполяризации).
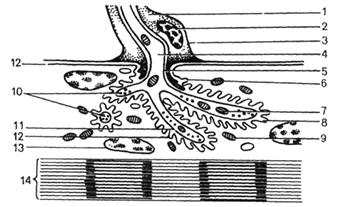
Рис. 56. Ультрамикроскопическое
строение нервно-мышечного окончания (схема).
1 - цитоплазма нейролеммоцита; 2 - ядро нейролеммоцита; 3 - плазмолемма нейролеммо-цита;
4 - осевой цилиндр нервного волокна; 5 - аксолемма; 6
- постсинаптическая мембрана (сарколемма); 7 - митохондрии в аксоплазме; 8 - синаптическая
щель; 9 - митохондрии мышечного волокна; 10 - пресинаптические
пузырьки; 11 - пресинаптическая мембрана (аксолемма); 12 - сарколемма; 13 - ядро мышечного волокна;
14 - миофибрилла.
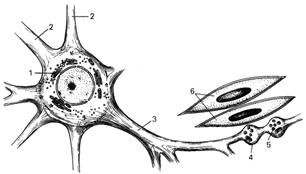
Рис. 57. Двигательные
нервные окончания в гладкой мышечной ткани.
1 - тело (перикарион) мультиполярного
нейрона; 2 - дендриты; 3 - аксон; 4 - варикозы
(утолщения) с синаптическими пузырьками; 5 - синаптические пузырьки; 6 - гладкомышечные клетки.
антитела против ацетилхолиновых рецепторов сарколеммы. Эти антитела
связываются с холинорецепторами постсинаптической
мембраны и инактивируют их, нарушая нервно-мышечное взаимодействие.
Инактивированные рецепторы подвергаются эндоцитозу и
замещаются новыми, которые ожидает та же участь.
Постсинаптическая мембрана моторного нервного окончания
содержит фермент ацетилхолинэстеразу, разрушающий
медиатор и ограничивающий этим срок его действия. Нарушения в нервно-мышечных
окончаниях вызывают развитие неизлечимого заболевания – миастения,
характеризующегося прогрессирующей мышечной слабостью и часто заканчивающегося
параличом дыхательной мускулатуры (межреберных мышц и диафрагмы). У больных в кови циркулируют антиацетилхолиновые
антитела.
Двигательные нервные окончания в
гладкой мышечной ткани
представляют собой четкообразные утолщения (варикозы)
нервного волокна, идущего среди неисчерченных гладких миоцитов (рис. 57). Варикозы
содержат адренергические или холинергические пресинаптические
пузырьки. Нейролеммоциты в области варикозов часто отсутствуют, и волокно проходит
"обнаженным".
Сходное строение имеют секреторные нервные
окончания (нейрожелезистые). Они представляют
собой концевые утолщения терминалей или утолщения по
ходу нервного волокна, содержащие пресинаптические
пузырьки, главным образом холинергические.
Рецепторные нервные окончания. Эти нервные окончания - рецепторы - рассеяны по всему
организму и воспринимают различные раздражения как из
внешней среды, так и от внутренних органов. Соответственно выделяют две большие
группы рецепторов: экстерорецепторы и интерорецепторы. К
экстерорецепторам (внешним) относятся слуховые, зрительные, обонятельные,
вкусовые и осязательные рецепторы. К интерорецепторам (внутренним) относятся висцеро-рецепторы (сигнализирующие о состоянии внутренних
органов) и вестибуло-проприорецепторы (рецепторы
опорно-двигательного аппарата). В зависимости от специфичности раздражения,
воспринимаемого данным видом рецептора, все чувствительные
окончания делят на механорецепторы, барорецепторы, хеморецепторы, терморецепторы и др.
По особенностям строения чувствительные окончания
подразделяют на свободные нервные окончания, т.е. состоящие только из
конечных ветвлений осевого цилиндра, и несвободные, содержащие в своем
составе все компоненты нервного волокна, а именно ветвления осевого цилиндра и
клетки глии. Несвободные окончания, кроме того, могут
быть покрыты соединительнотканной капсулой, и тогда они называются инкапсулированными.
Несвободные нервные окончания, не имеющие соединительнотканной капсулы,
называются неинкапсулированными (рис. 58).
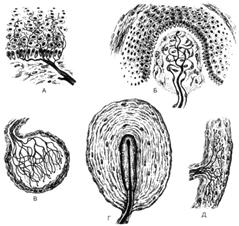
Рис. 58. Рецепторные
нервные окончания (схема по Р.В.Кристичу с
изменениями). А - свободные нервные окончания (боль); Б -
тельце Мейсснера (прикосновение); В - колба Краузе (холод); Г - тельце Фатера-Пачини
(давление); Д - тельце Руффини (тепло).
Свободные нервные окончания обычно воспринимают холод, тепло и боль. Такие
окончания характерны для эпителия. В этом случае миелиновые нервные волокна
подходят к эпителиальному пласту, теряют миелин, а осевые цилиндры проникают в
эпителий и распадаются там между клетками на тонкие терминальные ветви.
Очень разнообразны рецепторы в соединительной ткани.
Огромное большинство их представляет собой разной степени сложности ветвления
осевого цилиндра. В состав таких концевых аппаратов, как правило, входят нейролеммоциты, которые сопровождают все ветвления волокна
(это несвободные неинкапсулированные рецепторы)
(рис. 58).
Инкапсулированные рецепторы соединительной ткани при всем их разнообразии всегда
состоят из ветвления осевого цилиндра и глиальных клеток. Снаружи такие
рецепторы покрыты соединительнотканной капсулой. Примером
подобных окончаний могут служить весьма распространенные у человека пластинчатые
тельца - тельца Фатера-Пачини). Их
размеры 0,5×1-2 мм. В центре такого тельца располагается
внутренняя луковица, или колба, образованная видоизмененными
леммоцитами (рис. 59).
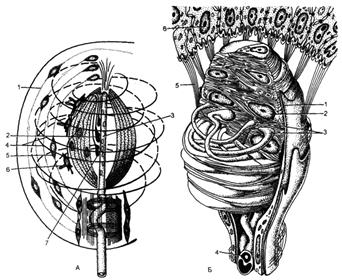
Рис. 59. Строение
инкапсулированных нервных окончаний.
А - пластинчатое тельце Фатера-Пачини:
1 - слоистая капсула; 2 - внутренняя луковица; 3 - дендрит чувствительной
нервной клетки; 4 - спиральные коллагеновые волокна; 5 - фиброциты; 6 - вторичночувствующие клетки с ресничками; 7 - синаптические контакты аксонов вторично чувствующих клеток
с дендритами чувствительной нервной клетки (по А.А.Отелину,
В.Р.Машанскому, А.С.Миркину);
Б - осязательное тельце: 1 - капсула;
2 -специальные клетки; 3 - нервные терминали; 4 -
миелиновое нервное волокно; 5 - опорные (поддерживающие) фибриллы; 6 - эпителий
(по Р.Кристичу с изменениями).
Миелиновое чувствительное нервное волокно теряет около
пластинчатого тельца миелиновый слой, проникает во внутреннюю луковицу и
разветвляется. Снаружи тельце окружено слоистой капсулой,
состоящей из фибробластов и спирально ориентированных волокон. Заполненные
жидкостью пространства между пластинками содержат коллагеновые микрофибриллы. Давление на капсулу передается через
заполненные жидкостью пространства между пластинками на внутреннюю луковицу и
воспринимается безмиелиновыми волокнами во внутренней
луковице. Пластинчатые тельца воспринимают давление и вибрацию. Они
присутствуют в глубоких слоях дермы (особенно в коже пальцев), в брыжейке и
внутренних органах.
К чувствительным
инкапсулированным окончаниям относятся осязательные тельца - тельца
Мейснера. Эти структуры овоидной
формы размерами 50-150×60 мкм. Они располагаются в верхушках
соединительнотканных сосочков кожи. Осязательные тельца состоят из
видоизмененных нейролеммоцитов - тактильных
клеток, расположенных перпендикулярно длинной оси тельца. Части тактильных
клеток, содержащие ядра, расположены на периферии, а уплощенные части,
обращенные к центру, формируют пластинчатые отростки, интердигитирующие
с отростками противоположной стороны. Тельце окружено тонкой капсулой.
Миелиновое нервное волокно входит в основание тельца снизу, теряет миелиновый
слой и формирует ветви, извивающиеся между тактильными клетками. Коллагеновые микрофибриллы и волокна связывают тактильные клетки с
капсулой, а капсулу с базальным слоем эпидермиса, так что любое смещение
эпидермиса передается на осязательное тельце.
К инкапсулированным нервным окончаниям относятся также
рецепторы мышц и сухожилий: нервно-мышечные веретена и нервно-сухожильные
веретена.
Нервно-мышечные веретена являются сенсорными органами в скелетных мышцах,
которые функционируют как рецептор на растяжение. Веретено состоит из
нескольких исчерченных мышечных волокон, заключенных в растяжимую
соединительнотканную капсулу, - интрафузальных
волокон.
1. Понятие о рефлекторной дуге
Нервная ткань входит в состав нервной системы,
функционирующей по рефлекторному принципу, морфологическим субстратом которого
является рефлекторная дуга. Рефлекторная дуга представляет собой цепь
нейронов, связанных друг с другом синапсами и обеспечивающих проведение
нервного импульса от рецептора чувствительного нейрона до эфферентного
окончания в рабочем органе.
Самая простая рефлекторная дуга состоит из двух
нейронов - чувствительного и
двигательного (рис. 60).
В подавляющем большинстве случаев между
чувствительными и двигательными нейронами включены вставочные,
или ассоциативные, нейроны. У высших животных
рефлекторные дуги состоят обычно из многих нейронов и имеют значительно более
сложное строение, чем на приведенном рисунке. Конкретные нервные связи будут
рассмотрены на примере коры головного мозга и мозжечка.
|
|
|
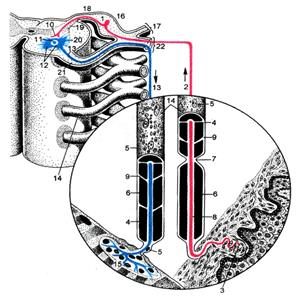
Рис. 60. Простая рефлекторная дуга (схема по В.Г.Елисееву, Ю.И.Афанасьеву, Е.Ф.Котовскому).
1
- чувствительная нервная клетка; 2 - дендрит чувствительной клетки; 3 -
рецептор в коже; 4 - плазмолемма нейролеммоцита; 5 -
ядра нейролеммоцитов; 6 - миелиновый слой; 7 -
узловой перехват нервного волокна; 8 - осевой цилиндр; 9 - насечка миелина; 10
-аксон чувствительной клетки; 11 - двигательная клетка; 12 - дендриты
двигательной клетки; 13 - аксон двигательной клетки; 14 - миелиновые волокна;
15 - эффектор на мышце; 16 - спинномозговой узел; 17 - дорсальная ветвь спинномозгового
нерва; 18 - задний корешок; 19 - задний рог; 20 - передний рог; 21 - передний
корешок; 22 - вентральная ветвь спинномозгового нерва.
2. Гистогенез
нервной ткани
Нервная ткань развивается из дорсальной эктодермы. У
18-дневного эмбриона человека эктодерма по средней линии спины дифференцируется
и утолщается, формируя нервную пластинку, латеральные
края которой приподнимаются, образуя нервные валики, а между валиками
формируется нервный желобок. Передний конец нервной пластинки
расширяется, образуя позднее головной мозг. Латеральные края продолжают
подниматься и растут медиально, пока не встретятся и не сольются по средней
линии в нервную трубку, которая отделяется от лежащей над ней эпидермальной эктодермы. Полость нервной трубки сохраняется
у взрослых в виде системы желудочков головного мозга и центрального канала
спинного мозга. Часть клеток нервной пластинки не входит в состав нервной
трубки и эпидермальной эктодермы и образует скопления
по бокам от нервной трубки, которые сливаются в рыхлый тяж, располагающийся
между нервной трубкой и эпидермальной эктодермой, - нервный
гребень (ганглиозная пластинка). Из нервной трубки в дальнейшем
формируются нейроны и макроглия центральной нервной
системы. Нервный гребень дает начало нейронам чувствительных (сенсорных) и
автономных ганглиев, клеткам мягкой мозговой и паутинной оболочек мозга и
некоторым видам глии: шванновским
клеткам, клеткам-сателлитам ганглиев, клеткам мозгового вещества надпочечников,
меланоцитам кожи, части клеток APUD-системы,
сенсорным клеткам каротидных телец и др.
В формировании ганглиев V, VII, IX и X черепных нервов
принимают участие, кроме нервного гребня, также нейральные
(нейрогенные) плакоды, представляющие собой
утолщения эктодермы по бокам формирующейся нервной трубки в краниальном отделе
зародыша.
Нервная трубка на ранних стадиях эмбриогенеза
представляет собой многорядный нейроэпителий,
состоящий из вентрикулярных или нейроэпителиальных
клеток. В дальнейшем в нервной трубке дифференцируется 4 концентрических
зоны: вентрикулярная (эпендимная),
субвентрикулярная, промежуточная (плащевая) и краевая
(маргинальная).
Вентрикулярная (эпендимная)
зона состоит из делящихся клеток цилиндрической формы. Ядро вентрикулярной
клетки мигрирует в люменальный конец клетки,
обращенной к центральному каналу. Клетки делятся и после деления ядра дочерних
клеток также мигрируют в апикальные части образующихся клеток, где происходит
репликация ДНК. Митотический цикл и цикл ядерной миграции продолжаются от 5 до
24 ч. Вентрикулярные (или матричные) клетки являются
предшественниками нейронов и клеток макроглии. Микроглия развивается из другого источника (см. ниже).
Предшественники глиальных клеток отличаются по присутствию глиального
фибриллярного кислого белка промежуточных филаментов
в делящихся клетках вентрикулярной зоны.
Субвентрикулярная зона состоит из клеток,
утративших способность к перемещению ядер, но сохраняющих высокую
пролиферативную активность. Субвентрикулярная зона
существует в области спинного мозга в течение нескольких дней, но в тех областях
головного мозга, где гистогенез совершается особенно интенсивно, формируются субвентрикулярные и экстравентрикулярные
герминативные
(камбиальные) зоны, существующие длительное время.
Так, экстравентрикулярная камбиальная зона мозжечка
исчезает у человека к 20 мес
постнатального онтогенеза.
Промежуточная (плащевая, мантийная) зона
состоит из клеток, переместившихся из вентрикулярной
и субвентрикулярной зон -нейробластов и глиобластов. Нейробласты
утрачивают способность к делению и в дальнейшем дифференцируются в нейроны. Глиобласты продолжают делиться и дают начало астроцитам и олигодендроцитам.
Способность к делению не утрачивают полностью и зрелые
астроциты, и олигодендроциты.
Новообразование нейронов прекращается в раннем постнатальном периоде. Поскольку
число нейронов в головном мозге составляет примерно 1 триллион, очевидно, в
среднем в течение всего пренатального периода в 1 мин
формируется 2 500 000 нейронов. Из клеток плащевого слоя образуются серое
вещество спинного и часть серого вещества головного мозга.
Маргинальная зона (краевая вуаль) формируется из
врастающих в нее аксонов нейробластов и макроглии и дает начало белому веществу. В некоторых
областях головного мозга клетки плащевого слоя мигрируют дальше, образуя
кортикальные пластинки - скопления клеток, из которых формируется кора большого
мозга и мозжечка.
По мере
дифференцировки нейробласта изменяется
субмикроскопическое строение его ядра и цитоплазмы. В ядре возникают участки
различной электронной плотности в виде мелких зерен и нитей. В цитоплазме
выявляются в большом количестве канальцы и цистерны гранулярной
эндоплазматической сети, уменьшается количество свободных рибосом и полисом,
значительного развития достигает аппарат Гольджи.
Специфическим признаком начавшейся специализации нервных клеток следует считать
появление в их цитоплазме тонких фибрилл - пучков нейрофиламентов
и микротрубочек. Количество нейрофиламентов,
содержащих белок - нейрофиламентный триплет, в
процессе специализации увеличивается. Тело нейробласта
постепенно приобретает грушевидную форму, а от его заостренного конца начинает
развиваться отросток - аксон (нейрит). Позднее дифференцируются другие
отростки - дендриты. Нейробласты
превращаются в зрелые нервные клетки - нейроны. Между нейронами
устанавливаются контакты (синапсы).
В процессе дифференцировки нейронов из нейробластов различают до-медиаторный
и медиаторный периоды. Для домедиаторного
периода характерно постепенное развитие в теле нейробласта
органелл синтеза - свободных рибосом, а затем эндоплазматической сети. В медиатор-ном периоде у юных нейронов появляются первые пузырьки,
содержащие медиатор, а в дифференцирующихся и зрелых нейронах отмечаются
значительное развитие органелл синтеза и секреции (гранулярная
эндоплазматическая сеть, аппарат Гольджи), накопление
медиаторов и поступление их в аксон, образование синапсов. Несмотря на
то, что формирование нервной системы завершается в первые годы постнатального
развития, известная пластичность центральной нервной системы сохраняется до
старости. Эта пластичность может выражаться в появлении новых терминален и новых синаптических
связей. Нейроны центральной нервной системы млекопитающих способны формировать
новые ветви (аксональное почкование) и новые синапсы (синаптическое
замещение). Пластичность проявляется в наибольшей степени в первые годы
после рождения, но частично сохраняется и у взрослых - при изменении уровней
гормонов, обучении новым навыкам, травме и других воздействиях. Хотя нейроны
постоянны, их синаптические связи могут
модифицироваться в течение всей жизни, что может выражаться, в частности, в
увеличении или уменьшении их числа. Пластичность при малых повреждениях мозга
проявляется в частичном восстановлении функций.
В популяции нейронов, начиная с ранних стадий развития
нервной системы и в течение всего онтогенеза, имеет место массовая гибель
клеток, достигающая 25-75 % всей популяции. Эта запрограммированная
физиологическая гибель клеток (апоптоз) наблюдается
как в центральной, так и в периферической нервной системе; при этом мозг теряет
около 0,1 % нейронов. У человека ежегодно погибает около 10 млн
нервных клеток.