Тема 4. СОБСТВЕННО СОЕДИНИТЕЛЬНЫЕ ТКАНИ
План:
1. Общая
характеристика и классификация соединительных тканей.
2. Рыхлая
волокнистая соединительная ткань.
3. Плотная
волокнистая соединительная ткань.
4. Соединительные
ткани со специальными свойствами.
5. Гистогенез
и возрастные особенности соединительной ткани.
1. Общая характеристика и
классификация
соединительных тканей
В понятие
«соединительные ткани» (ткани внутренней среды, опорно трофические ткани)
объединяются ткани, развивающиеся из единого источника – мезенхимы и имеющие общую
соединительно-трофическую функцию.
Общие
принципы организации соединительных тканей:
1)
внутреннее
расположение в организме;
2)
преобладание
межклеточного вещества над клетками;
3) межклеточное вещество делится на волокнистое и аморфное;
4)
многообразие
клеточных форм;
5)
общий
источник происхождения – мезенхима.
Главными компонентами соединительных тканей являются волокна - производные клеток, коллагенового и эластического типов и основное (аморфное) вещество, играющее роль интегративно-буферной метаболической среды, а также и клеточные элементы. Клетки здесь создают и поддерживают количественное и качественное соотношение состава неклеточных компонентов.
Органная специфичность клеточных элементов соединительной
ткани выражается в количестве, форме и соотношении различных видов клеток, их
метаболизме и функциях, оптимально приспособленных к функции органа.
Специфика соединительной
ткани обнаруживается и в соотношении клеток и неклеточных структур в различных участках
тела.
Межклеточное
вещество соединительной ткани
Основное (или
аморфное) вещество состоит из белков и углеводов. Белки представлены в
основном коллагеном, а также альбу минами и глобулинами. Углеводы представлены полимерными формами,
в основном гликозаминогликанами (сульфатированными – хондроитинсерными
кислотами, дерматансульфатом и др.) Углеводные компоненты удерживают воду, в
зависимости от содержания воды ткань может быть более или менее плотной.
Аморфное вещество обеспечивает транспорт
веществ из крови клеткам и обратно, в том числе транспорт из соединительной
ткани в эпителиальную.
Оно образуется за счет деятельности
прежде всего фибробластов – коллагенов и гликозаминогликанов, а так же за счет веществ
плазмы крови – альбуминов и глобулинов. Если в
нем меньше воды, то оно более плотное и наоборот.
Волокнистое вещество представлено коллагеновыми, эластическими
и ретикулярными волокнами. В различных органах соотношение названных волокон
неодинаково: в рыхлой волокнистой соединительной ткани преобладают коллагеновые
волокна.
Каждое коллагеновое
волокно состоит из двух химических компонентов:
1) фибриллярного белка
коллагена;
2)
углеводного
компонента – гликозаминогликанов и протеогликанов.
Оба данных компонента синтезируются
фибробластами и выделяются во внеклеточную среду, где и осуществляется их
сборка и построение волокна. Коллаген образован тремя полипептидными α-цепями, которые, скручиваясь (имеет спиральное
строение, образуют молекулу тропоколлагена. Коллагеновые волокна толщиной 1–20
мкм – это изогнутые тяжи, состоящие из фибрилл с
поперечной исчерченностью. Они обладают высокой прочностью и малой
растяжимостью.
В структурной организации
коллагенового волокна выделяют пять уровней.
I уровень – полипептидный. Коллаген представлен
полипептидными цепочками, состоящие из трех
аминокислот – пролина, глицина, лизина.
II уровень – молекулярный, представлен молекулой белка
коллагена длиной 280 нм, шириной 1,4 нм, состоящей из трех полипептидных
цепочек, закрученных в спираль.
III уровень – протофибриллярный (толщина
10 нм, состоит из нескольких продольно расположенных молекул коллагена,
соединенных между собой водородными связями).
IV уровень – микрофибриллы (толщиной от 11–12нм,
И более). Они состоят из 5-6протофибрилл, связанных
боковыми связями.
V уровень – фибрилла (или коллагеновое волокно)
толщина 1–10мкм, состоящее из нескольких
микрофибрилл – в зависимости от толщины,
связанных гликозаминогликанами и протеогликанами. Коллагеновые волокна имеют
поперечную исчерченность, обусловленную как расположением аминокислот в
полипептид ной цепи, так и расположением цепей в молекуле коллагена.
Коллагеновые волокна с помощью углеводных компонентов соединяются в пучки
толщиной до 150 мкм.
В
зависимости
от порядка расположения аминокислот в полипептидных цепочках, от степени их
гидроксилирования и от качества углеводного компонента различают двенадцать
типов белка коллагена, из которых хорошо изучены только пять типов.
Эти
разновидности белка коллагена входят в состав не только коллагеновых волокон,
но и в состав базальных мембран эпителиальной ткани и сосудов, хрящевых тканей,
стекловидного тела и других
образований. При развитии некоторых патологических процессов происходит распад
коллагена и поступление его в кровь. В плазме крови биохимически определяется
тип коллагена, следовательно, определяется и предположительная область его
распада и его интенсивность.
Эластические
волокна толщиной от 3 до 10 мкм образованы белком эластином,
который также синтезируется фибробластами. В отличие от коллагеновых
эластические волокна способны растягиваться в 1,5 раза, после чего возвращаются
в исходное состояние. Эластические волокна анастомозируют и переплетаются между
собой, образуя сети, окончатые пластины и мембраны. Эластические волокна
характеризуются высокой эластичностью, способностью растягиваться и
сокращаться, но незначительной прочностью.
Они тоньше коллагеновых, не имеют поперечной исчерченности, по ходу разветвляются и
анастомозируют друг с другом, образуя эластическую сеть. Химический состав
эластических волокон – белок эластин и гликопротеины. Оба компонента
синтезируются и выделяются фибробластами, а в стенке сосудов – гладкомышечными клетками. Белок эластин отличается от
белка коллагена как составом аминокислот, так и их
гидроксилированностью. Структурно эластическое волокно организовано следующим
образом: центральная часть волокна представлена аморфным компонентом из молекул
эластина, а периферическая часть –
мелкофибриллярной сетью. Соотношение аморфного и фибриллярного компонента в эластических волокнах может быть
различным. В большинстве волокон преобладает аморфный компонент. При равенстве
аморфного и фибриллярного компонентов волокна называют элауниновыми.
Встречаются также окситалоновые эластические волокна, состоящие только из
фибриллярного компонента. Локализуются эластические волокна, прежде всего в тех
органах, которые постоянно изменяют свой объем – в легких, сосудах.
Ретикулярные
волокна – тонкие (от 100 нм до 1,0 мкм), разветвленные,
малорастяжимые, переплетаясь между собой, образуют сеть, в ячейках которой
расположены клетки. Эти волокна образуют
строму лимфоидных органов (органов
кроветворения и иммунной системы), печени, поджелудочной железы и других
паренхиматозных органов, окружают капилляры, кровеносные и лимфатические
сосуды, а также связаны с ретикулярными клетками.
Ретикулярные
волокна по своему составу близки к коллагеновым волокнам. Ретикулярные волокна
состоят из коллагена третьего типа и углеводного
компонента. Они тоньше коллагеновых, имеют слабо выраженную
поперечную исчерченность. Разветвляясь и анастомозируя, они образуют мелкопетлистые сети, откуда и происходит их название. В ретикулярных
волокнах в отличие от коллагеновых более выражен углеводный компонент, который
хорошо выявляется солями азотнокислого серебра, поэтому эти волокна называют
еще аргирофильными. Следует помнить, что аргирофильными свойствами обладают и
незрелые коллагеновые волокна, состоящие из белка преколлагена. По своим
физическим свойствам ретикулярные волокна занимают промежуточное положение
между коллагеновыми и эластическими. Они образуются за
счет деятельности ретикулярных клеток. Локализуются в основном в кроветворных
органах, составляя их строму.
Функции соединительных
тканей:
1)
трофическая
(метаболическая);
2)
опорная;
3)
защитная
(механическая и имунная);
4)
структурная (пластическая);
5)
терморегуляторная;
6)
регенеративная;
7) запасающая и др.
Классификация
соединительных тканей. Виды
соединительной ткани различаются между собой составом и соотношением клеток,
волокон, а также физико-химическими свойствами аморфного межклеточного
вещества. Соединительные ткани подразделяются на собственно соединительную
ткань (волокнистые соединительные ткани и соединительные ткани со
специальными свойствами) и скелетные ткани. Разновидности скелетных
ткане: три вида хрящевой ткани (гиалиновая, эластическая, волокнистая), две
вида костной ткани (фиброзно-волокнистая и пластинчатая), а также цемент и
дентин зуба (рис. 25).
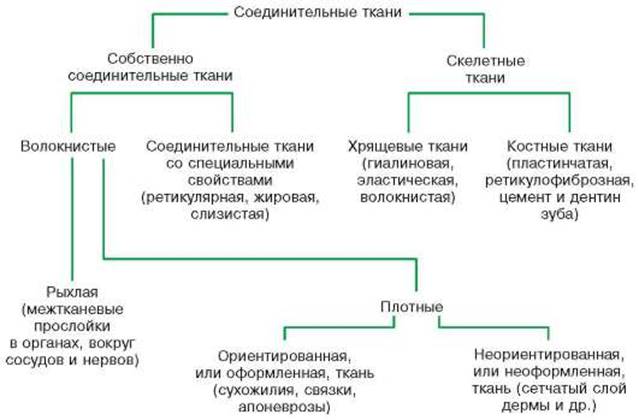
Рис. 25.
Классификация соединительных тканей.
1.
Рыхлая
волокнистая соединительная ткань
Наиболее
распространенными в организме являются собственно соединительная волокнистая ткань
и особенно рыхлая волокнистая неоформленная ткань, которая входит в состав
практически всех органов, образуя строму, слои и прослойки, сопровождая
кровеносные сосуды.
Она
располагается преимущественно по ходу кровеносных и лимфатических сосудов, нервов,
покрывает мышцы, образует строму (каркас) большинства внутренних органов,
собственную пластинку слизистой оболочки, подсерозную основу, адвентициальную
оболочку.
Морфо-функциональная
характеристика рыхлой волокнистой
неоформленной соединительной ткани. Состоит из клеток и межклеточного
вещества, которое продуцируют в основном фибробласты. Межклеточный матрикс включает аморфное и волокнистое вещество. По мере
старения фибробласты превращаются в многоотростчатые фиброциты, которые
образуют трехмерную сеть, в пространствах которой располагаются различные
клетки (рис. 26).
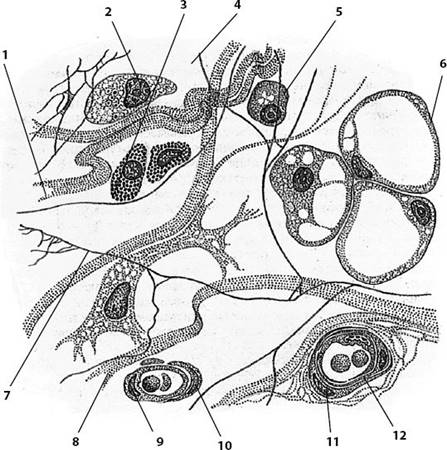
Рис. 26.
Рыхлая волокнистая соединительная ткань: 1 - коллагеновое волокно; 2 -
макрофагоцит; 3 - тканевый базофил; 4 - аморфное межклеточное (основное) вещество;
5 - плазмоцит (плазматическая клетка); 6 - адипоцит (жировая клетка); 7 -
эластическое волокно; 8 - фибробласт; 9 - эндотелиоцит; 10 - перицит; 11 -
миоцит; 12 - кровеносный сосуд.
Рыхлая
волокнистая соединительную ткань отличается от других разновидностей
соединительных тканей:
1) многообразие клеточных форм (девять
клеточных типов);
2)
преобладание
в межклеточном веществе аморфного вещества над волокнами.
Функции рыхлой волокнистой соединительной
ткани:
1)
трофическая;
2)
опорная (образует строму паренхиматозных
органов);
3)
защитная
(неспецифическая и специфическая защита, участие в иммунных реакциях);
4)
депо
воды, липидов, витаминов, гормонов;
5)
репаративная
(пластическая).
Типы клеток (клеточные популяции) рыхлой волокнистой
соединительной ткани:
1)
фибробласты;
2)
макрофаги
(гистиоциты);
3)
тканевые
базофилы (тучные клетки);
4)
плазмоциты;
5)
жировые
клетки (адипоциты или липоциты);
6)
пигментные
клетки;
7)
адвентициальные
плетки;
8)
перициты;
9)
клетки
крови — лейкоциты (лимфоциты, нейтрофилы).
Фибробласты — преобладающая популяция клеток этой
ткани, главные тканеобразующие клетки, так как отвечают за синтез и выделении в межклеточную среду
белков коллагена и эластина, а также гликозаминогликанов. Они неоднородны по степени
зрелости и функциям, потому делятся на:
1)
малодифференцированные
клетки;
2)
дифференцированные
(или зрелые клетки, или собственно фибробласты);
3)
старые
фибробласты (дефинитивные);
4)
миофибробласты;
5)
фиброкласты.
Преобладающей
формой являются зрелые фибробласты, у них развиты зернистая
ЭПС и комплекс Гольджи, остальные органеллы развиты слабо (рис. 27). В фиброцитах
зернистая ЭПС и пластинчатый комплекс редуцированы. В
цитоплазме фибробластов содержатся микрофиламенты, содержащие сократительные
белки актин и миозин, но особенно развиты эти органеллы в миофибробластах,
благодаря которым они осуществляют стягивание молодой соединительной ткани при
образовании рубца. У фиброкластов в цитоплазме большого
количество лизосом, они выделяют в межклеточную среду ферменты для расщепления
коллагеновых или эластических волокон на фрагменты, а затем фагоцитируют
расщепленные фрагменты. То есть, фиброкласты отвечают за лизис лизиса
межклеточного вещества, в том числе волокон (например, при инволюции матки после
родов).
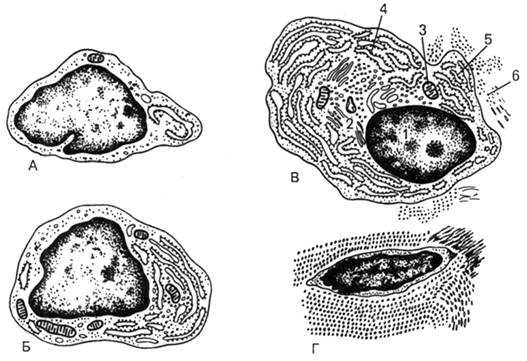
Рис. 27.
Ультрамикроскопическое строение фибробласта на разных стадиях дифференцировки
(схема по Н.А.Юриной и А.И.Радостиной, с изменениями).
А - малодифферецированный; Б - молодой; В - зрелый; Г -
фиброцит; 1 - ядро; 2 -аппарат Гольджи; 3 - митохондрии; 4 - рибосомы и
полирибосомы; 5 - гранулярная эн-доплазматическая сеть; 6 - коллагеновые
фибриллы.
Таким образом,
различные формы фиброкластов образуют межклеточное вещество соединительной
ткани (фибробласты), поддерживают его в определенном структурном и
функциональном состоянии (фиброциты), разрушают его при определенных условиях
(фиброкласты). Благодаря этим свойствам фибробластов осуществляется
репаративная функция соединительной ткани.

Рис. 28. Происхождение и структура системы мононуклеарных
фагоцитов (СМФ).
Макрофаги (макрофагоциты). В 1882 г. И.И.
Мечников впервые описал фагоцитоз, был удостоен Нобелевской
премии. После двух-трех суток циркуляции моноцит покидает кровеносное русло. За
сутки у человека обменивается 0,5–1,0×109 моноцитов,
причем количество их в циркулирующей крови примерно в 20 раз меньше, чем в
тканях.
В 70-х годах XX века
сформировалось представление о системе мононуклеарных фагоцитов (СМФ),
включающей группу клеток, объединенных общностью происхождения из моноцитов
крови (которые, в свою очередь, имеют общего предшественника стволовую
кроветворную клетку), строения и функции – активный фагоцитоз и пиноцитоз (рис. 28, 29).
А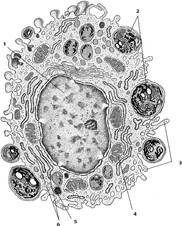 Б
Б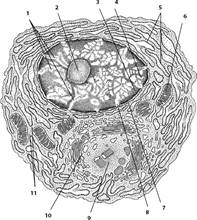
В Г
Г 
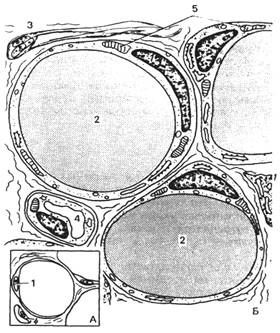
Рис. 29. Клетки рыхлой соединительной ткани.
А
- макрофаг (по Крстичу, с
изм.): 1 – ядро; 2 – фаголизосомы (вторичные лизосомы); 3 – псевдоподии; 4 –
митохондрия; 5 – остаточное тельце; 6 – цистерны гранулярной эндоплазматической
сети.
Б -. плазматическая клетка (по Крстичу, с изм.): 1 – радиально
ориентированные скопления хроматина («спицы»); 2 – ядрышко; 3 – гетерохроматин;
4 – ядро; 5 – гранулярная эндоплазматическая сеть; 6 – митохондрия; 7 – ядерная
мембрана; 8 – пора в ядерной оболочке; 9 – клеточный центр; 10 – комплекс
Гольджи; 11 – свободные рибосомы.
В - ретикулярная
клетка (по Крстичу, с
изм.): 1 – отростки; 2 – ретикулярные волокна; 3 – лизосомы; 4 – отростки; 5 –
ундулирующая мембрана; 6 – ядро; 7 – митохондрия; 8 – комплекс Гольджи; 9 –
гранулярная эндоплазматическая сеть. Г
- перицит (по Крстичу, с изм.): 1 – базальная мембрана; 2 – нервное
волокно; 3 – тело перицита; 4 – отростки; 5 – нервное волокно; 6 – тело
перицита; 7 – эндотелиоцит; 8 – синаптические пузырьки; 9 – базальная мембрана;
10 – капилляр.
Особенностью макрофагов
является большое количество различных функциональных форм лизосом. Макрофаги
секретируют большое количество различных биологически активных веществ. Зрелые
макрофаги не способны делиться.
В то же время СМФ является
саморегулируемой системой. В ответ на раздражение периферического звена
(тканевые макрофаги), усиливается деление (пролиферация) клеток-предшественниц
моноцитов в костном мозге, увеличивается количество моноцитов костного мозга и
крови, они мигрируют в ткани и превращение в макрофаги.
Основные
функции макрофагов – это участие в естественном, специфическом,
противоопухолевом иммунитете и секреция различных биологически активных
веществ. Они осуществляют
защитную функцию, прежде всего посредством фагоцитоза крупных частиц. По
современным данным макрофаги являются полифункциональными клетками. Образуются
макрофаги из моноцитов после их выхода из кровеносного русла. Макрофаги
характеризуются структурной и функциональной гетерогенностью в зависимости от
степени зрелости, области локализации, а также от их активации антигенами или
лимфоцитами.
Защитная функция
макрофагов проявляется в разных формах:
1)
неспецифическая
защита (посредством фагоцитоза экзогенных и эндогенных частиц и их
внутриклеточного переваривания);
2)
выделение
во внеклеточную среду лизосомальных ферментов и других веществ;
3)
специфическая
(иммунологическая защита).
Макрофаги
подразделяются на фиксированные и свободные. Макрофаги соединительной ткани
являются подвижными или блуждающими и называются гистиоцитами (тканевые макрофаги).
Различают макрофаги
серозных полостей (перитонеальные и плевральные), альвеолярные,
макрофаги печени (купферовские клетки), макрофаги центральной нервной системы — глиальные
макрофаги, остеокласты.
Все виды
макрофагов объединяются в мононуклеарную фагоцитарную или макрофагическую
систему организма.
По
функциональному состоянию макрофаги подразделяются на резидуальные (неактивные)
и активированные. В зависимости от этого отличается и их внутриклеточное
строение.
Наиболее
характерной структурной особенностью макрофагов является содержание множества лизосом и фагосом.
Особенностью гистоцитов является наличие на их поверхности
многочисленных складок, инвагинаций и псевдоподий, отражающих передвижение
клеток или захват ими разнообразных частиц. В плазмолемме макрофагов содержатся
разнообразные рецепторы, с помощью которых они распознают различные, в том
числе и антигенные частицы, а также разнообразные биологически активные
вещества.
Фагоцитируя
антигенные вещества, макрофаги выделяют, концентрируют, а затем выносят на
плазмолемму их активные химические группировки – антигенные детерминанты, а затем передают
их на лимфоциты. Данная функция называется антигенпредставляющей. С помощью
данной функции макрофаги запускают антигенные реакции, так как установлено, что
большинство антигенных веществ не способно запускать иммунные реакции
самостоятельно, т. е. действовать непосредственно на рецепторы лимфоцитов.
Кроме того, активированные макрофаги выделяют некоторые биологически активные
вещества – монокины, которые регулирующее
влияние на различные стороны иммунных реакций.
Макрофаги
принимают участие в заключительных стадиях иммунных реакций как гуморального,
так и клеточного иммунитета. В гуморальном иммунитете они фагоцитируют иммунные
комплексы «антиген – антитело», а в клеточном иммунитете под влиянием
лимфокинов макрофаги приобретают киллерные свойства и могут разрушать
чужеродные, в том числе и опухолевые, клетки.
Таким образом,
макрофаги не являются иммунными клетками, но принимают участие в иммунных
реакциях. Они также синтезируют и выделяют в межклеточную среду около сто
различных биологически активных веществ. Поэтому макрофаги можно отнести к
секреторным клеткам.
Тканевые базофилы (тучные клетки) являются истинными клетками рыхлой
волокнистой соединительной ткани. Функция: регуляция местного тканевого
гомеостаза. Это достигается посредством синтеза тканевыми базофилами и
последующим выделением в межклеточную среду гликозаминогликанов (гепарина и
хондроитинсерных кислот), гистамина, серотонина и других биологически активных
веществ, которые оказывают влияние на клетки и межклеточное вещество
соединительной ткани.
Наибольшее
влияние эти биологически активные вещества оказывают на микроциркуляторное
русло, где вызывают повышение проницаемости гемокапилляров, усиливают
гидратацию межклеточного вещества. Продукты тучных клеток оказывают влияние на
иммунные реакции и на процессы воспаления и аллергии.
Источники
образования тучных клеток в настоящее время окончательно не установлены.
Для ультраструктурной
организации тканевых базофилов характерно наличие в цитоплазме двух типов
гранул:
1) метахроматических гранул,
окрашивающихся основными красителями с изменением цвета окраски;
2) ортохроматических гранул,
окрашивающихся основными красителями без изменения цвета и представляющих собой
лизосомы.
При возбуждении
тканевых базофилов из них выделяются биологически активные вещества следующими
способами:
1) с помощью выделения гранул –
дегрануляцией;
2) с помощью диффузного
выделение через мембрану гистамина, который повышает сосудистую проницаемость и
вызывает гидратацию
основного вещества, усиливая тем самым воспалительную реакцию.
Тучные клетки
принимают участие в иммунных реакциях. При попадании в организм некоторых
чужеродных веществ плазмоциты синтезируют иммуноглобулины класса Е, которые затем адсорбируются на цитолемме тучных клеток.
При повторном попадании в организм этих же антигенов на поверхности тучных
клеток образуются иммунные комплексы «антиген — антитело», которые вызывают резкую дегрануляцию тканевых базофилов, а выделяющиеся в
большом количестве биологически активные вещества обусловливают быстрое
наступление аллергических и анафилактических реакций.
Плазматические клетки (плазмоциты) являются клетками иммунной системы
(эффекторные клетки гуморального иммунитета).
Образуются
плазмоциты из В-лимфоцитов при воздействии на них антигенных веществ.
Большинство их
локализуется вблизи мелких кровеносных сосудов в лимфоидных органах -
лимфоузлах, селезенке, миндалинах, фолликулах, в слизистой оболочке
пищеварительной и дыхательной систем органах иммунной системы, но значительная
часть плазмоцитов распределяется в соединительной ткани.
Функции
плазмоцитов –
синтез и выделение в межклеточную среду антител – иммуноглобулинов, которые подразделяются на пять классов.
В плазмоцитах
хорошо развит синтетический и выделительный аппарат. На электроннограммах
плазмоцитов видно, что почти вся цитоплазма заполнена зернистой
эндоплазматической сетью, кроме небольшого участка, который примыкает к ядру и
в котором расположен пластинчатый комплекс Гольджи и клеточный центр. При
изучении плазмоцитов под световым микроскопом при обычной гистологической
окраске –
гематоксилин эозин, они имеют округлую или овальную форму, базофильную
цитоплазму, эксцентрично расположенное ядро, содержащее глыбки гетерохроматина
в виде треугольников (колесообразное ядро). К ядру прилежит бледно окрашенный
участок цитоплазмы – «светлый дворик», в
котором локализуется комплекс Гольджи. Число плазмоцитов отражает интенсивность
иммунных реакций.
Жировые клетки (адипоциты). Различают два типа жировой ткани: белую
и бурую, которые сформированы соответственно белыми или бурыми адипоцитами
(рис. 29, 30).
Клетки белого жира – крупные, шаровидные,
диаметром 50–120 мкм. Как видно на рисунках 30, 31 в адипотцитах белого жира в
цитоплазме крупная липидная капля оттесняет ядро к плазмолемме, органоидов мало. В адипоцитах бурого жира
липидные капли мелкие и многочисленные, ядро в центре цитоплазмы, митохондрий очень
много, также много лизосом.
Бурый цвет жировым клеткам
придают железосодержащие пигменты – цитохромы митохондрий.
Функции
жировых клеток:
1)
синтез и внутриклеточное накопление липидов; (адипоциты поглощают жирные кислоты из
капиллярной крови и синтезируют из них жиры). Бурые адипоциты содержат мелкие
капельки жира и очень много митохондрий, за счет обилия цитохромов – бурый
цвет. Бурого жира много у новорожденных, у взрослого почти весь жир – белый.
2) депо энергии;
2)
депо
воды;
3)
депо
жирорастворимых витаминов,
4)
теплоизоляция (белый жир),
5)
теплообразование (бурый жир).
|
|
Рис. 30. Строение белой жировой
ткани (схема по Ю.И.Афанасьеву). |
|
|
|
Рис. 31. Строение бурой жировой
ткани (схема по Ю.И.Афанасьеву). |
|
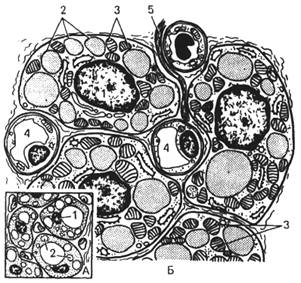
Пигментные клетки (пигментоциты, меланоциты) – клетки отростчатой формы, содержащие в цитоплазме пигмент меланин. В их цитоплазме большое количество меланосом (гранул меланина) размером 15-25 нм и рибосом. Часть меланосом из меланоцитов мигрирует в кератиноциты шиповатого и базального слоев эпидермиса. В цитоплазме меланоцитов содержатся также биологически активные амины, которые могут принимать участие вместе с тучными клетками в регуляции тонуса стенок сосудов.
Пигментные
клетки не являются истинными клетками соединительной ткани, так как, во-первых,
они локализуются не только в соединительной, но и в эпителиальной ткани,
залегают в эпидермисе, особенно наружных половых органов, в радужке и
собственно сосудистой оболочке глазного яблока, в мягкой мозговой оболочке, а
во вторых, они образуются не из мезенхимальных клеток, а из нейробластов
нервных гребешков. На
1 мм2 поверхности кожи
приходится 1200-1500 меланоцитов. У представителей черной и желтой рас
количество их значительно больше. «Цвет» глаз зависит от генетически
детерминированного количества меланоцитов в радужке. Чем меньше их, тем
светлее радужка.
Адвентициальные клетки – малоспециализированные
клетки, сопровождающие кровеносные сосуды, то есть локализуются в
адвентиции сосудов. Они имеют вытянутую и уплощенную форму. Цитоплазма данных
клеток слабобазофильна и содержит незначительное количество органелл. Одни
авторы рассматривают адвентициальные клетки как самостоятельные клеточные
элементы соединительной ткани, другие считают, что они являются источником для
развития фибробластов, жировых и гладкомышечных клеток.
Перициты – отростчатые клетки,
локализующиеся снаружи стенок
кровеносных капилляров, окружают
их и соприкасаются отростками с эндотелиальной клеткой, то есть входящие
в состав стенки капилляров. Они передают последним
нервное возбуждение, что способствует накоплению или потере клеткой жидкости.
Результат этого – расширение или сужение
просвета капилляра.
Ретикулярная клетка –
удлиненная клетка, ее многочисленные отростки, которые соединяются и образуют
сеть, а в ее в углублениях располагаются ретикулиновые волокна. Ретикулярные
клетки образуют строму органов иммунной системы и кроветворения. При внедрении
инородных частиц, инфекции ретикулоциты округляются, отделяются от ретикулярных
волокон и становятся способными к фагоцитозу. Ретикулярные клетки образуют
строму органов иммунной системы и кроветворения.
В соединительной ткани
находятся также лимфоциты и зернистые лейкоциты.
В норме в соединительной
ткани обязательно содержатся в различных количествах клетки крови – лимфоциты и нейтрофилы.
При воспалительных состояниях количество их резко увеличивается.
Разнообразные клетки соединительной ткани
функционально связаны в единую систему благодаря многочисленным факторам
взаимодействия, особенно в процессах воспаления и посттравматической
регенерации, при нарушении водно-солевого режима организма и др.
2.
Плотная волокнистая соединительная ткань
Плотная
волокнистая соединительная ткань характеризуется сильным развитием волокон
межклеточного вещества. Если в рыхлой
волокнистой соединительной ткани превалируют клетки и аморфное вещество над
волокнами, а в плотной, наоборот, основную массу соединительной ткани
составляют волокна. В межклеточном веществе
преобладают пучки коллагеновых волокон. Количество клеток (фиброцитов)
незначительное.
В
зависимости от характера расположения волокон плотная волокнистая
соединительная ткань подразделяется на оформленную (волокна данного вида ткани
располагаются упорядоченно, чаще всего параллельно друг другу) и неоформленную
(волокна располагаются беспорядочно).
Плотная оформленная волокнистая
соединительная ткань формирует сухожилия, связки, фасции, фиброзных
мембраны. Эластическая плотная оформленная соединительная ткань образует стенку
артерий эластического типа, эластический конус гортани и ее голосовые связки,
желтые связки, выйную связку копытных. Главными элементами ее являются тесно
прилежащие друг к другу эластические волокна, между которыми залегают малочисленные
фиброциты.
Плотная
волокнистая неоформленная соединительная ткань образует сетчатый слой дермы
кожи. Помимо содержания большого числа волокон, плотная волокнистая
соединительная ткань характеризуется бедностью клеточных элементов, которые
представлены в основном фиброцитами.
Строение
сухожилия. Сухожилие состоит в основном из плотной оформленной
соединительной ткани, но содержит также и рыхлую волокнистую соединительную
ткань, образующую прослойки.
На поперечном и
продольном разрезе сухожилия видно, что оно состоит из параллельно
расположенных коллагеновых волокон, образующих пучки I, II и III порядков.
Пучки I порядка – наиболее тонкие, отделены
друг от друга фиброцитами. Пучки II порядка состоят из нескольких пучков I
порядка, окруженных по периферии прослойкой рыхлой волокнистой соединительной
ткани, составляющей эндотеноний. Пучки III порядка состоят из пучков II порядка
и окружены более выраженными прослойками рыхлой волокнистой соединительной
ткани – перитенонием.
Все сухожилие по периферии окружено эпитенонием.
В прослойках
рыхлой волокнистой соединительной ткани проходят сосуды и нервы, обеспечивающие
трофику и иннервацию сухожилия.
4. Соединительные ткани
со специальными свойствами
Соединительные ткани со специальными
свойствами расположены лишь в определенных органах и
участках тела и характеризуются особыми чертами строения и своеобразной
функцией (жировая, ретикулярная, пигментная).
Жировая
ткань выполняет
трофическую, депонирующую, формообразующую и терморегулирующую функции. Жировая ткань
подразделяется на два типа: белую, образованную однокапельными жировыми
клетками, и бурую, образованную многокапельными. Группы жировых клеток
объединены в дольки, отделенные друг от друга перегородками рыхлой волокнистой неоформленной
соединительной ткани, в которой проходят сосуды и нервы. В свою очередь, каждая
жировая клетка окутана сетью коллагеновых и ретикулярных волокон, в петлях
которой проходят многочисленные капилляры и залегают соединительнотканные (в
том числе тучные) клетки. У человека преобладает белая жировая ткань, часть
ее окружает органы, сохраняя их положение (например, почки, лимфатические узлы,
глазное яблоко и др.); заполняет пространства еще не функционирующих органов
(например, молочная железа); замещает красный костный мозг в эпифизах длинных
трубчатых костей. При похудании жировая ткань, выполняющая указанные функции,
меняется относительно мало. Большая часть жировой ткани является резервной
(подкожная основа, сальники, брыжейки, жировые подвески толстой кишки,
субсерозная основа).
Функции
белой жировой ткани:
1) депо энергии;
2) депо воды;
3) депо жирорастворимых
витаминов;
4) механическая защита
некоторых органов (глазного яблока и др.);
5) теплоизоляция.
Функция
бурой жировой ткани: теплообразование.
То есть, белый и бурый жир по-разному участвуют в терморегуляции организма.
Бурая жировая ткань у человека содержится
в небольшом количестве, имеется главным образом у новорожденного ребенка. Она
расположена в области шеи, за грудиной, около лопаток, в подмышечной ямке,
вдоль позвоночника, в окружности подключичной артерии, под кожей спины и
боковых поверхностей туловища, в средостении и брыжейках.
Подобно белому жиру, бурая жировая ткань также
сформирована дольками, образованными многокапельными жировыми клетками. Бурый
цвет, как было выше сказано, обусловлен
обилием митохондрий с цитохромами. По
сравнению с клетками белой жировой ткани в них значительно больше митохондрий.
Окислительная способность бурых жировых клеток примерно в 20 раз выше белых и
почти в 2 раза превышает окислительную способность мышцы сердца. При понижении
температуры окружающей среды повышается активность окислительных процессов в
бурой жировой ткани. При этом выделяется тепловая энергия, обогревающая кровь в
кровеносных капиллярах. В регуляции теплообмена определенную роль играют
симпатическая нервная система и гормоны мозгового вещества надпочечников –
адреналин и норадреналин, которые стимулируют активность тканевой липазы,
расщепляющей триглицериды на глицерин и жирные кислоты. Это приводит к
высвобождению тепловой энергии, обогревающей кровь, протекающую в
многочисленных капиллярах между липоцитами. При голодании бурая жировая ткань
изменяется меньше, чем белая.
Бурая жировая ткань поддерживает
температуру тела животных во время зимней спячки и температуру новорожденных
детей. Основная функция бурой жировой ткани заключается в теплообразовании, так
как окисление и фосфорилирование в их митохондриях разобщены.
Слизистая соединительная ткань встречается только у зародыша в
провизорных органах и прежде всего в составе пупочного канатика. Она состоит в
основном из межклеточного вещества, в котором локализуются фибробластоподобные
клетки, синтезирующие муцин (слизь).
Клеточные элементы здесь представлены гетерогенной группой клеток, дифференцирующихся из мезенхимных клеток на протяжении эмбрионального периода. Эти клетки составляют дифферон: фибробласты – миофибробласты – гладкие мышечные клетки. Они отличаются способностью к синтезу виментина, десмина, актина, миозина. Слизистая соединительная ткань пупочного канатика ("вартонов студень") синтезирует коллаген IV типа, характерный для базальных мембран, ламинин, гепаринсульфат. Между клетками этой ткани в первой половине беременности в большом количестве обнаруживается гиалуроновая кислота, что обусловливает желеобразную консистенцию основного вещества. Фибробласты студенистой соединительной ткани слабо синтезируют фибриллярные белки. Лишь на поздних стадиях развития зародыша в студенистом веществе появляются рыхло расположенные коллагеновые фибриллы.
Пигментная соединительная ткань представляет собой участки ткани, в
которых содержится скопление меланоцитов в (область сосков, мошонки, анального отверстия,
сосудистая оболочка глазного яблока).
5. Гистогенез и возрастные особенности соединительной
ткани
Различают эмбриональный и постэмбриональный гистогенез
соединительных тканей. В процессе эмбрионального
гистогенеза мезенхима приобретает черты тканевого строения раньше закладки
других тканей. Этот процесс в различных органах и системах происходит
неодинаково и зависит от их неодинаковой физиологической значимости на
различных этапах эмбриогенеза.
В дифференцировке мезенхимы отмечаются топографическая
асинхронность как в зародыше, так и во внезародышевых
органах, высокие темпы размножения клеток, волокнообразования, перестройка
ткани в процессе эмбриогенеза – резорбция путем апоптоза и новообразование
ткани.
Постэмбриональный гистогенез в нормальных физиологических условиях происходит
медленнее и направлен на поддержание тканевого гомеостаза, пролиферацию
малодифференцированных клеток и замену ими отмирающих клеток. Существенную роль
в этих процессах играют межклеточные внутритканевые взаимодействия,
индуцирующие и ингибирующие факторы (интегрины, межклеточные адгезивные
факторы, функциональные нагрузки, гормоны, оксигенация, наличие
малодифференцированных клеток).
У новорожденных
и детей в волокнистой соединительной ткани, в аморфном веществе содержится много
воды, связанной гликозоаминогликанами. Коллагеновые волокна тонкие и состоят не
только из белка, но и из преколлагена. Эластические
волок на хорошо развиты. Аморфный и волокнистые
компоненты соединительной ткани в совокупности обусловливают эластичность и
упругость кожи у детей. С увеличением возраста в постнатальном онтогенезе
содержание гликозаминогликанов в аморфном веществе ткани уменьшается, а
соответственно снижается и содержание воды. Коллагеновые волокна разрастаются и
образуют толстые и грубые пучки. Эластические волокна в значительной степени
разрушаются. Вследствие этого кожа у пожилых и старых людей становится
неэластичной и дряблой.